薛建辉主编
森林生态学
中国林业出版社2006年第一版
第3章 种群及其基本特征
第4章 林木种内与种间关系
第3章 种群及其基本特征
【本章提要】种群具有许多不同于个体的群体特征,如出生率、死亡率、增长率、年龄结构、性比、空间分布格局。本章通过对种群数量动态过程、种群调节以及生态对策的剖析,使学生在种群层次上更进一步地了解生物与环境的相互关系。
种群生态学(population ecology)是研究生物种群与环境之间相互关系的科学。种群不仅是构成物种的基本单位,而且也是构成群落的基本单位。
种群是人类利用与保护或控制生物物种的对象。因此,种群生态学与生态环境建设和物种保护有着密切的关系,涉及珍贵、稀有和濒危物种的保护与开发,以及有害生物的控制。
3.1种群的基本概念
在自然界中,生物很少以孤立的个体形式长期存在,它或多或少、直接或间接地依赖别的生物而存在。生物也只有形成一个群体才能繁衍后代。因此,个体必须依赖群体而存在.群体则是个体发展的必然结果。
种群(population)是同一物种占有一定空间和一定时间的个体集合群。种群这个术语在生物学科中广泛应用,除生态学外,在进化论、遗传学和生物地理学中也经常使用。
种群虽是由许多个体集合而成,但并不是个体的简单组合。种群具有自己独特的性质、结构,特别是具有自动调节的能力,以适应空间和时间上的变化。因此,种群既反映了构成它的个体的特性,也反映了它构成群落的特性。种群的研究既有助于个体研究的深化,又是群落及生态系统研究的基础。
种群是物种(species)具体的存在单位、繁殖单位和进化单位。一个物种通常可以包括许多种群,不同种群之间存在着明显的地理隔离,长期隔离的结果有可能发展为不同的生态种(ecospecies),甚至产生新的物种。如油松从河南、山东向北分布到辽宁、内蒙古,其分布比较广阔,显然不能说它们是一个种群,可能因地理隔离、人为作用、生境分化等影响,在种内形成不同的类群。它们在形态上、生理上或生态习性上分别表现出显著的差异,并随着生态环境的长期特化逐渐显现出变异现象。所以,物种的进化是通过种群表现出来的,种群亦是物种进化的单位。 ,
事实上,种群的空间界限和时间界限并不是十分明确的,除非种群栖息地具有清楚的边界,如岛屿、湖泊等。因此,种群的空间界限常掌由研究者根据调查的目的予以划定。如油松种群可以抽象地泛指森林中的全部油松林,也可以具体指森林中一小块油松林。生长在不同地段内的同种各个集合体,可以理解为一个种群,也可以理解为彼此独立的种群。这样,种群可以作为抽象的概念在理论上加以应用如种群生态学、种群遗传学理论和种群研究方法,也可以作为具体的研究对象又分为自然种群和实验种群,实验种群指实验室内饲养或培养的一群生物。
种群可以由单体生物(unitary organism)或构件生物(modular organism)组成。在由单体生物组成的种群中,每一个体都是由一个受精卵直接发育而来,个体的形态和发育都可以预测,如哺乳类、鸟类、两栖类和昆虫都是单体生物的例子。相反,由构件生物组成的种群,受精卵首先发育成一结构单位或构件,然后发育成更多的构件。发育成分支结构的形式和时间是不可预测的,大多数植物,水绵和珊瑚是构件生物。高等植物通过积累构件而生长。通常构件包括叶子、芽和茎,花也是一种类型的构件。一些构件生物,如树木和海扇,主要垂直生长,而有根状茎的草和结硬壳的海绵沿基质侧面扩散生长。构件生物各部分之间的连接可能会死亡和腐烂,这样就形成了许多分离的个体,这些个体来自于同一个受精卵,并且基因型相同,这样的个体被称为无性系分株(ramets)。
种群除了与组成种群的个体具有类似的生物学特性外,如生长、分化(性别等)、进化、死亡等,还具有个体所不具备的群体特性,例如出生率、死亡率、平均寿命、年龄结构和性比等。这些都说明了种群的整体性和统一性。概括地说,作为群体属性,自然种群应具有以下三个主要特征:①空间特征,即种群有一定的分布区域和分布方式;②数量特征,即种群具有一定的密度、出生率、死亡率、年龄结构和性比;③遗传特征,即种群具有一定基因组成,即系一个基因库,以区别其他物种,但基因组成具有随着时间改变其遗传特性的能力,即进化、适应能力。
种群由个体组成,但具有自己独立的特征、结构和功能,种群是组成群落和生态系统的基本成分。种群生态学就以生物种群及其环境为研究对象,研究这些群体属性,包括种群的基本特征、数量动态及调节规律,林木种内与种间关系。种群生态学的主要任务是研究生物种群的数量结构变化及变化的原因,要定量地研究种群的出生率、死亡率、迁入率和迁出率,以便能够了解是什么因素影响着种群渡动的范围及种群发生变化的规律,了解种群波动所围绕平均密度的变化以及了解种群衰落和绝灭的原因。了解的目的是为了能够控制种群。与种群生态学有密切关系的种群遗传学(population genetics) 研究种群的遗传过程,包括遗传变异、选择、基因流、突变和遗传漂变等。20世纪60年代,很多生物学家认识到研究种群生态学和种群遗传学的局限性,发现种群个体数量和遗传特性动态有着密切的关系,并力图将两个独立的分支学科有机的整合起来,从而提出了种群生物学。生态遗传学和进化生态学就是在这种思想影响下迅速发展起来的。进化理论的基本原理之一是自然选择作用与生物个体,而种群通过自然选择而进化,因此,种群生态学与种群遗传学有着密切的关系。特别是今年来随着分子生物学的渗透而于1992年诞生的分子生态学的发展,对种群生物学问题的研究有了较大进展。
自然条件下,任何种群都不是孤立存在,而是与生态系统中的其它生物有密切关系,为了研究的方便,分析研究对象限定为单种种群。
3.2种群的基本特征
种群虽然是由个体组成的,但种群具有个体所不具有的特征。从个体到种群有一个质的飞跃,在群体水平上,表现出新的特征,即种群具有个体所不具备的各种群体特征。
3.2.1 种群密度
种群密度(density)是指单位面积或体积内种群的个体数量,也可以是生物量或能量。在调查分析种群密度时,首先应区别单体生物和构件生物。因为个体数只能反映单体生物的种群大小,对构件生物就必须进行两个层次的数量统计,即合子产生的个体数(它与单体生物的个体数相当)以及组成每个个体的构件数。由于生物的多样性,具体数量统计方法随生物种类或栖息地条件而异。密度通常代表单位面积(或空间)上的个体数目,但也有用每片叶子、每个植株、每个宿主为单位的,此外,如果研究者对进化个体(evolutionary individual)的数量感兴趣,就应当考虑无性系的数量。许多植物都是无性繁殖,个体本身就是一个无性系的“种群”,因此研究植物种群动态,必须重视个体水平以下的构件组成“种群”的意义,这是植物种群与动物种群的重要区别。
种群密度的统计首先就是划分研究种群的边界,森林呈大面积连续分布,种群边界不明显,所以在实际工作中往往需要研究者根据需要自己确定种群边界。森林中种群个体大小或经济价值相差悬殊,为经营上的方便.需要分层统计种群密度.如森林经营中,林分密度一般仅指检尺直径以上的林木种群密度,不包括幼苗和幼树的密度,有时在森林更新调查时,单独统计幼苗密度和幼树密度,分析和评价森林的更新状况。
林木种群密度统计是建立在样地(标准地)的基础上,例如在某一样地(面积为0.25hm²������。)油松的数量为100株,可以推出谈区油松的种群密度为400株/hm²������。样方必须具有代表性,并通过随机取样来保证结果可靠,同时用数理统计方法估计变异和显著性。但森林中下木和草本植物因是丛生多分枝或个体矮小,不易查数,通常不以单位面积上植株个体数计量种群密度,多采用多度(调查样地上个体的数目多少)或盖度(植物枝叶覆盖地面的百分数)反映种群的密度,它们只能采用目测估计法.填写调查的种群属于何等级。因为森林中下木和草本植物的种类和数量对森林更新有很大影响。
在种群生态学上。密度不是按种的分布区计算,而是依据其在分布区内最适生长的空间计算,这种密度称为生态密度(ecological density),而把全分布区内的平均密度称为粗密度(crude density)以示区别。如一片面积为l0hm²������的马尾松林,林木总株数为30 000株,但其中有2hm²������的面积为裸露的岩石,2 hm²������ 的水域面积。因此,实际分布有马尾松林的面积只有6 hm²������。则该马尾松林的粗密度为3 000株/hm²������、生态密度为5 000株/hm²������。
当进行森林群落分析时,还应了解各种植物种群的相对数量,即种群的相对密度。相对密度(relative density)是指一个种群的株数占样地内所有种群总株数的百分比。例如样地上有50株林木,其中30株是红松种群,那么红松的相对密度是30/50=0.60或60%。
种群密度一方面反映了种群的数量或大小,另一方面反映了种群个体所占有的空间面积,它关系到种群对光能(能流)和营养物质(物流)利用效率,直接影响到种群及群落的生产力,是提高单位面积产量的关键因素之一。林学家常把森林的管理和林地质量评价建立在林木密度调查的基础上。种群密度过大时,每一种生物都会以特有的方式作出反应,如森林自然稀疏,另一方面,种群密度也有一个最低限度,种群密度过低时,使种群的异性个体不能正常相遇和繁殖,会引起种群灭亡,表现出产量过低。但是种群密度的上限主要是由生物的大小和该生物所处的营养级而决定的。一般来说,生物越小,单位面积中的个体数量就越多,例如1 hm²������ 森林中,可容纳幼树的数量就比大树的数量多。生物所处的营养级越低,种群的密度也就越大,例如同样是l hm²������的森林,其中植物的数量比食草动物多,而食草动物的数量又比食肉动物多。因此,合理密植在农业、林业生产实践中具有重要的意义。
影响种群大小或密度的因素主要有:
①种群的繁殖特性:各种生物的繁殖力不同,如微生物几个小时可繁殖数代;草本植物一般1~3年结实一次,乔木可能几年才结实一次。
②种群的结构:如一个种群内不同年龄的个体或不同性别的个体比例。
③种内和种间的关系:即种内遗传变异和物种自然选择等。
④物理环境因子:如光照、温度、水分、土壤、大气和火等。种群密度的高低在多数情况下取决于环境中可利用的物质和能量的多少。当环境中拥有可利用的物质和能量最丰富、环境条件最适应时,某种群可达到该环境下的最大密度,这个密度称为“饱和点”。维持种群最佳生长状况的密度称为最适密度。最大密度和最适密度是林业经营、作物栽培、动物饲养、鱼类养殖应首先考虑的问题,也是人类自身生存所必须考虑的问题。密度是种群特征的一个重要参数。密度决定着种群的能流、资源的可利用性、种群内部生理压力的大小以及种群的散布和种群的生产力。
3.2.2 种群的空间结构
种群内由于生境的多样性,以及种内个体之间的竞争,每一种群在一定空间都会呈现出特定的分布形式。种群内个体在生存空间的分布方式或配置特点,称为种群的空间分布格局(spatial pattern)或内分布型(internal distribution pattern )。它是由种群的生物学特性,种内、种间关系和环境因素的综合影响所决定的。
3.2.2.1 种群的分布格局类型
种群个体的空间分布格局大致可分为3种类型:均匀型(uniform)、随机型(random)和集群(成群)型(clumped)(图3-1)。 (1)随机分布
(1)随机分布
每一个个体在种群领域中各个点上出现的机会是相等的,并且某一个体的存在不影响其他个体的分布。随机分布在自然界不很常见,只有当环境均一、种群个体间没有彼此吸引或排斥的情况下,才能出现随机分布。用种子繁殖的植物在初入侵到一个新的地点时,常呈随机分布。
(2)均匀分布
种群内个体在空间上是等距离分布形式。均匀分布是由于种群内个体间的竞争所引起的,例如森林中植物为竞争阳光(林冠)和土壤中营养物(根际),沙漠中植物为竞争水分。干燥地区所特有的自毒(autotoxin)现象,以阻止同种植物籽苗的生长是导致均匀分布的另一个原因。人工林属于均匀分布,但它是由于人为均匀栽植而形成的。
(3)集群分布
种群内个体在空间的分布极不均匀,常成群、成簇、成块或呈斑点状密集分布,这种分布格局即为集群分布,也叫成群分布和聚群分布。集群分布是自然界最常见的内分类型。
集群分布形成的原因是:①种群的繁殖特点。从母树上散布种子,落在附近,种子长成植株,形成群状;有些植物果实内含有多粒种子,如松树球果,这些种子长成植株总是非常靠近,是簇状;植物的无性繁殖,形成密集的团聚:;如伐根萌芽和根蘖形成的集群。②资源分布不均匀造成环境条件的差异。如天然过熟林中,老树因枯死风倒或雷击等原因所形成的林中空地(林窗)中更新起来的幼苗、幼树;局部地形的微起伏和土壤条件的局部差异。③种内种间的相互作用。它们之间有可能是直接的有利作用,或间接的互为环境。④动物对植物种子的收集和贮藏能导致植物的集群生长。
种群个体的集群分布是对自然的长期选择和适应的结果,它有许多有利的方面:①具有保护作用。在恶劣的环境如大风和高温中,植物能相互保护。如果正好处于树木线之下的森林被大面积砍伐,则这一地区的许多成熟林常常很难更新,因为风、夏季高温以及冬季低温的共同影响,使得天然更新或人工造林非常困难。②利于繁殖。许多物种的种群大小和密度必须超过一个低限,才能繁殖和生存。③增加个体间的基因交流,丰富遗传多样性。群聚促成种群的遗传变异和多态性,遗传多样性增加了种群内不断的变化和难以预测的环境中的生存能力。有助于形成宽广的地理分布区和该种的种内竞争,如果种群太小,近亲繁殖会降低遗传多样性,种群绝灭的危险性增加。④有益的种内竞争。虽然种内竞争(同种不同个体之闻的竞争)有不利的一面,但它常常使种群保持旺盛生长,并能很好地适应其物理环境。有病的和适应能力差的个体在竞争中会被淘汰。通过在种内选择高适应性的个体,这种竞争能使该种群在种间(不同种的个体之间)竞争中处于有利的地位。
种群个体的集群分布也有不利的方面:①加剧种内竞争。由于自然选择会减少种间竞争,因此个体在与其同种不同个体的竞争中所遭受的不利影响,常常比与其他种的个体的竞争中受到的不利影响更严重。正如达尔文(1859)所说:“同种不同个体之间的竞争永远是最激烈的,因为它们常活动于同一区域.需要同样的食物,遭受同样的危脸”,种群内个体过分拥挤能导致多种竞争。林分密度大的森林中的林木,为伸展枝条所需的空间而竞争,为光照、土壤水分和土壤养分而竞争,为所需的根系空间而竞争。干旱环境中过密的森林,其整个种群的生长势可能低下,因为其中任何个体都不能得到足够的资源以战胜其相邻个体。②导致环境恶化。由于种内个体密度的不断增加,在高密度下,没有哪个个体可得到足够的营养。对资源利用的普遍重叠程度,导致环境恶化,结果是急剧地减少个体数。③疾病的传播。寄生物、捕食动物以及病原生物,一般以其寄生或被捕食动物的高密度聚集中获益不少,被攻击者越多,被攻击者之间的接触越频繁,就越能促使病原生物或寄生物的扩散和生长。因此,在混交林中,病虫害的传播速度常常比纯林中低得多。④个体间的相互干预。如果种群内个体过分密集.个体之间的接触非常频繁,林内树干挤压能造成树冠摩擦,损害形成层,影响林木个体的正常生长型。
同一种群的分布格局并不一定局限于某一种固定的形式,有些种群在不同的发育阶段可能有不同的分布格局。如当某一种群初时侵入,依靠种子自然撒播而呈随机分布;随后则因无性繁殖形成集群分布.最后可能因为竞争或其他原因又变为随机分布或均匀分布。因此.应对种群的自然属性与环境之间的关系进行全面地综合分析,才能对种群的分布格局和动态有较全面系统的认识。
3.2.2.2 种群分布格局的检验
(1)相邻个体最小距离法
种群空间格局的检验方法很多,如果种群的密度和个体间的最小距离能够精确测量,则可采用相邻个体最小距离(nearest-neighbor distance)法检验内分布型。
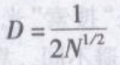
式中 D——个体间最小距离的理论值;
N——种群密度。
![]()
式中 d——个体间最小距离的观测值;
di——第i个随机选择的个体与相邻个体的最小距离
n—一观测次数。
![]()
式中 J一种群分布型的判别指标;
当J=1时即为随机分布;当J<1为集群分布;当J>1时为均匀分布。
(2)空间分布指数法
种群分布格局的检验还可以用空间分布指数(index of dispersion)法检验分布型。空间分布指数由方差/平均数比率即I=S2/![]() ≈0。如图3-1中的分布区分成许多小方块,进行样方取样和统计分析,如果个体是均匀分布,则各方格内个体数量相等的,方差应该等于零,所以S2/
≈0。如图3-1中的分布区分成许多小方块,进行样方取样和统计分析,如果个体是均匀分布,则各方格内个体数量相等的,方差应该等于零,所以S2/![]() ≈0;同样如果小方块中个体是随机分布,则样方中个体数出现频率将符合泊松分布序列S2/
≈0;同样如果小方块中个体是随机分布,则样方中个体数出现频率将符合泊松分布序列S2/![]() =1;如果个体是集群分布,则样方中含很少个体数的样本和含较多个体数的样本的出现频率将较泊松分布的期望值高,从而S2/
=1;如果个体是集群分布,则样方中含很少个体数的样本和含较多个体数的样本的出现频率将较泊松分布的期望值高,从而S2/![]() 的值明显大于1。因此,I=0属均匀分布;I=1属随机分布;I>1属集群分布。
的值明显大于1。因此,I=0属均匀分布;I=1属随机分布;I>1属集群分布。
其中
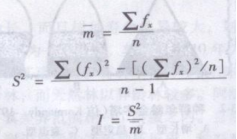
式中 x——样方中某种个体数;
f一一含z个体样方的出现频率;
n——样本总数。
集群分布又可进一步按种群本身的分布状况划分为均匀群、随机群和集群群,后者具有两级的集群分布。
内分布型的研究是静态研究,比较适合于植物、定居或不太活动的动物。构建生物的构件包括地面的枝条系统和地下的根系系统,其空间排列是一重要生态特征,对种群个体的适应和生存具有重要意义,例如在林冠层中,植物个体的枝叶系统的排列决定着光的摄取效率。同样土壤中根系分支的空间分布决定着水和营养物的获得。另外,枝叶系统是“搜索”光的,根系系统是“逃避”干旱的,这与动物依活动和行为进行搜索和逃避不同,靠的是控制构件生长的方向。
植物重复出现的构件的空间排列,可以称为建筑学结构(architecture),它是决定植物个体与环境相互关系和个体间相互作用的多层次的等级结构系统。构件建筑学结构的特征.主要视分支的角度、节间的长度和芽的死亡、休眠和产生新芽的概率。例如,草本植物可分为密集生长型和分散生长型两类。密集生长的草类.其节间短,营养枝聚集成簇,如生草、丛草类;分散生长型的草类。节间长,构件间相距较远,如车轴草。所以植物生态学应进一步强调个体和构件的空间排列。
3.2.3种群的年龄结构和性比
3.2.3.1种群的年龄结构
种群的年龄结构(age-structure)是指种群内个体的年龄分布状况,即各年龄或年龄组的个体数占整个种群个体总数的百分比结构。它是种群的重要特征之一。年龄结构直接关系到一个种群当前的出生率、死亡率和繁殖特点,对种群的未来发展有重要影响。了解种群的年龄结构,可以预测种群的发展趋势。研究种群的年龄结构对深入分析种群动态和进行预测预报具有重要价值。
(1)种群年龄结构的基本类型
一般用年龄金字塔(age pyramid)的形式来表示种群的年龄结构。年龄金字塔是以不同宽度的横柱从上到下配置而成的图(图3-2),横柱高低位置表示从幼年到老年的不同年龄组,宽度表示各年龄组的个体数或在种群中所占的百分比。按锥体形状,可划分为3个基本类型。
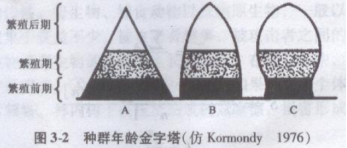
A.增长型 B.稳定型 C.褒退型
①增长型种群(Increasing population):锥体呈典型金字塔型,基部宽,顶部狭,表示种群中有大量幼体,而老年个体较少,种群的出生率大于死亡率,是迅速增长的种群。
②稳定型种群(stable population):锥体形状呈钟型,老中幼比例介于增长型和衰退型种群之间,种群的出生率和死亡率大致相平衡,即幼、中个体数大致相同,老年代表个体数较少,代表稳定型种群。
③衰退型种群(declining population):锥体呈壶型,基部比较狭,而顶部比较宽,种群中幼体比例减少而老体比例增大,种群的死亡率大于出生率,种群的数量趋于减少。
(2)种群年龄结构的基本生活时期
根据植物种群中个体的生长发育阶段,可以划分为以下几个基本生活时期:
①休眠期:植物以具有生活力的种子、果实或其他繁殖体(块根、地下茎)处于休眠状态之中。
②营养生长期:从繁殖发芽开始到生殖器官形成以前。这个时期还可细分为幼苗、幼年(幼树)、成年3个时期。
③生殖期:这一时期的特点是植物的营养体已基本定型,性器官成熟,开始开花结实,多年生多次结实的木本植物进入生殖期后.每年还要继续增加高度、粗度和新的枝叶,在每年的一定季节形成花、果和种子,但体形增长速度渐趋平缓。
④老年期:种群的个体到达老年期时.即使在生长良好的条件下,营养生长也很滞缓,繁殖能力逐渐消退,抗逆性减弱,植株逐渐趋向死亡。
这4个时期所占时问的长短,因植物种类不同而异,同一种类因所处生境条件的不同亦有差异。
在许多植物种类中,年龄结构仅能为种群提供有限的描述。因为其生长率是不可预测的,与年龄没有密切关联,一些植物可能比同种同龄的其他个体长得更大。在这些情况下,个体大小群(size classes)如生物量、覆盖面积或者树木胸高直径在生态学研究中可能比年龄更有效。
构件生物种群的年龄结构有个体年龄和组成个体的构件年龄两个层次。作为构件生物,植物体的年龄结构是由年轻的、正在生长发育和参与繁殖的部分与衰老的部分组成的,并且,叶、枝、根的活动性也随着年龄变化而变化。如果对所有叶子不加区别,就会忽略这样一个事实,其他生物如食草动物会对它们区别对待。
林木种群的个体寿命长,而且树种寿命差异较大,划分龄级组时间尺度不同,针叶树和硬阔叶树20年为一个龄级.软阔叶树10年为一个龄级,萌生起源和速生树种5年为一个龄级甚至更短。将林木种群年龄结构分为同龄林或异龄林结构.人工林多数为同龄林,而天然林以异龄林较多。同龄林是组成树木的年龄基本相同,如果有差异,其差异范围在一个龄级之内。自然条件下,同龄林一般是在裸地上,具有喜光、耐干旱的先锋树种所形成,种群难于连续更新,往往是上层林木衰老.林冠疏开(林窗),种群才得以更新,如桦木林和山杨林。异龄林是组成种群的林木年龄差异较大,有几个龄级的林木生存,由具有耐荫、喜湿的耐荫及中性树种所形成,林冠下可以正常更新,如云杉林、红松林。这样当裸地被先锋树种所占据,立地对该种群又是适宜,若不考虑其他种的种子来源,其该种群不断增长,成为增长型种群。喜光树种形成同龄林,到成熟后缺少同种更新幼苗幼树,常被其他树种所替代,将成为衰退型种群。异龄林种群能在林冠下自行更新,不断有幼树到达主林冠层,基本是稳定型种群。
3.2.3.2 性比 、
性比(sex ratio)指一个种群的所有个体或某年龄组的个体中雌性与雄性的个体数目的比例。性比是种群结构的重要特征之一,它对种群的发展有较大的影响,如果两性个体的数目相差过于悬殊,不利于种群的生殖。种群的性比同样关系到种群的出生率、死亡率和繁殖特点。大多数生物种群的性比接近1:1。植物中虽多数种是雌雄同株,没有性比问题,但某些雌雄异株植物如银杏、杨树等其性比可能变异较大,因此研究这些植物的性比具有重要意义。与性比相关联的因素,还有个体性成熟的年龄,也是影响种群繁殖的内在因素。
3.2.4种群的出生率与死亡率
3.2.4.1 出生率
出生率(natality)是指种群在单位时间内产生新个体数占总个体数的比率。出生是泛指生物产生新个体的过程,不管它是有性生殖(分娩、孵化或种子发芽),还是无性繁殖(分裂或分蘖等)。出生率有绝对出生率和相对出生率两种表示方法:
绝对出生率B=ΔNn/Δt
相对出生率b=ΔNn/(N•��������Δt)
式中 N-------种群的总个体数
ΔNn——新生的个体数;
Δt——时间增量。
一般的出生率是以相对出生率来表示,例如某一种现有个体数为1 000,在100天内,新出生的个体数为300,则每天平均生长率为:b=:300/1 000×100×100%=0.3%。此外.种群的出生率也可以用特定年龄出生率(age-specific natality)表示,即按不同的年龄组计算其出生率,不仅可以知道种群的出生率,而且可以知道整个不同年龄或年龄组在出生率上的差异。
出生率分为生理出生率(physiological natality)和生态出生率(ecological natality)。生理出生率又叫最大出生率(maximum natality),是指种群在理想条件下,无任何生态因子的限制,繁殖只受生理因素决定的最大出生率。对于某个特定种群来说,生理出生率是一个常数,当然完全理想的环境条件即使在实验室里也很难建立,它的意义在于是一个常数,以便实际的观察数作比较.因此在应用的时候,必须说明测定的条件。生态出生率又叫实际出生率(realized natality),是指在一定时期内,种群在特定环境条件下实际繁殖的个体数。
3.2.4.2死亡率
死亡率(mortality)代表一个种群的个体死亡情况。死亡率同出生率一样,也可以用特定年龄死亡率(age-specific mortality)表示,即按不同的年龄组计算。死亡率也可分为生理死亡率(physiological mortality)和生态死亡率(ecological mortality)。生理死亡率又叫最小死亡率(minimum mortality),是指在最适条件下个体因衰老而死亡,即每个个体都能活到该种群的生理寿命,因而使种群死亡率降到最低。对野生生物来说.生理死亡率同生理出生率一样是不可能实现的,它只有理论意义和比较意义。由于受环境条件、种群本身大小、年龄组成的影响以及种问的捕食、竞争等,实际死亡率远远大于理想死亡率。
生态死亡率是指在一定条件下的实际死亡率,可能有少数个体能话到生理寿命。最后死于衰老,但大部分个体将因饥饿、疾病、竞争、被寄生、恶劣的气候条件或意外事故等原因而死亡。
死亡是生物种群的一种必然现象,死亡率受环境条件、种群密度等因素的影响,环境条件恶劣,种群死亡率高,反之死亡率低。种群密度大,死亡率高,反之死亡率低。种间的竞争、捕食也是影响种群死亡率的最直接的主要原因。在农、林业生态系统之中,种群死亡率不仅受自然因素的影响,还受人为因索(输入、输出、干扰、调控)的影响。
除此以外,用存活率(survival rate)表示常比死亡率更有实用价值,生态学家对存活率更感兴趣,即存活个体数比死亡个体数更重要,假如用d表示死亡率,则存活率等于1-d。因此存活率通常吼生命期望(1ife expectancy)来表示,生命期望就是种群中某一特定年龄个体在未来所能存活的平均年数。
3.2.5生命表
把观测到的种群中不同年龄个体的存活数和死亡数编制成表,称为生命表(dynamic life table)。它反映了种群发展过程中从出生到死亡的动态变化,最早应用于人口统计(human demography),主要在人寿保险事业中,用来估计不同年龄组人口的期望寿命。由珀尔(Raymond Pear 1921)引人普通生物学。随着统计科学的发展和年龄鉴定技术的进步,生命表愈来愈广泛地应用于动植物的研究。
3.2.5.1动态生命表
动态生命表(dynamic life table)是根据对同年出生的所有个体存活数目进行动态监测的资料而编制的,这类生命表也称为同生群生命表(cohort 1ife table)。这种生命表对植物比较合适.因为植物固定不动,可以对每个个体进行标记和作图,通过跟踪观察一个特定个体群,如1hm2。样地上某树种幼苗的存活情况,直至全部死亡而建立起来的。但是乔木树种因其寿命很长,要做到连续跟踪记载难度较大。
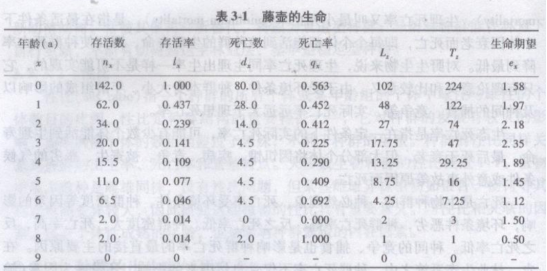
康内尔(Conell 1970)对某岛固着在岩石上的甲壳动物,生活在海滨潮间带的藤壶(Balanus glandula)从1959—1968年10年的精心观察编制出藤壶生命表(表3-1)。
其中,x:按年龄分阶段划分的单位时问期限,如年、月、日等;
nx:x期开始时的存活数;
lx:x期开始时的存活率;
dx:从x到x+1的死亡数;
引自Krebs,1978
qx:从x到x+l的死亡率;
ex:x期开始时的生命期望或平均余年。
Tx和Lx栏一般可不列入表中,但为计算气方便需要Tx和Lx栏。Lx是从x到x+1期的平均存活数,即Lx=(nx+nx+1)/2。Tx则是进入x龄期的全部个体在进入x期以后的存活体个体数总年数,即Tx=∑Lx,例如T0=L0+L1+L2+L3+…,T0=L0+L1+L2+L3+…,从这个生命表可获得三方面信息:
①存活曲线(survivorship curve):以lgnx,栏对x栏作图可得存活曲线。存活曲线直观地表达了该同生群的存活过程。Deevey(1947)曾将存活曲线分为3个类型(图3-3)。
I型:曲线凸型,表示在接近生理寿命前只有少数个体死亡,即几乎所有个体都能达到生理寿命。如大型兽类和人的存活曲线;藤壶、一些耐荫的阔叶树种的存活曲线接近这一类型。
Ⅱ型:曲线呈对角型,各年龄死亡数相等。许多鸟类、中性树种和一些耐荫树种的存活曲线接近这一类型。
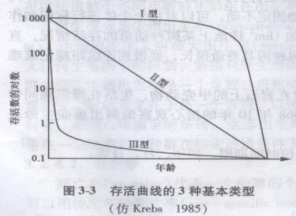
Ⅲ型:曲线凹型,幼年期死亡率较高。如鱼卵、真菌的孢予、喜光树种的存活曲线属于这一类型。
②死亡率曲线:以qx栏对x栏作图。藤壶在第一年死亡率很高,以后逐渐降低,接近老死时死亡率迅速上升。
③生命期望:ex表示该年龄期开始时平均能存活的年限,即预期还能活多少年。e0为种群的平均寿命。
3.2.5.2静态生命表
静态生命表(static life table)是根据某一种特定时间对种群作一年龄分布(结构)的调矗,它适用于世代重叠的生物,并掌握各年龄组的死亡率(数)再用统计学处理而编制的生命表。
静态生命表能够反映出种群出生率和死亡率随年龄而变化的规律,但却无法分析死亡的原因,也不能对种群密度制约过程的种群调节作定量分析。依据静态生命表,也难以建立更详细的种群模型,但它的优点是很容易使我们看出种群的生存对策和生殖对策,而且比较容易编制。
可根据种群普查时,如1hm2的某种林分在普查时的年龄分布进行调查分析统计。
动态生命表和静态生命表的关系可用图3-4来表示。图中纵坐标表示年龄,横坐标表示时间。连续追踪t0,t1,……时段中所繁殖的动植物就是动态生命表。因此,动态生命表也称为特定年龄生命表或水平生命表(age-specific or horizontal life table)。图中表示的根据t1时间所作年龄结构的生命表就属于静态生命表,也称特定时间生命表或垂直生命表(time-specific or vertical life table)。
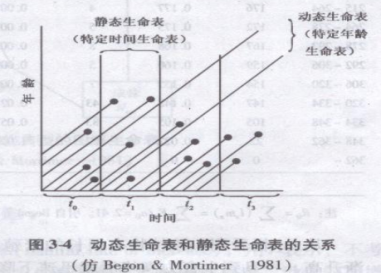
同生群生命表中个体经历了同样的环境条件,而静态生命表中个体出生于不同年(或其他时间单位),经历了不同的环境条件。因此编制静态生命表等于假定种群所经历的环境条件是年复一年没有变化的,但实际上并非如此。因此有的学者时静态生命表持怀疑态度,但在难以获得动态生命表数据时,如果应用得法,还是有价值的。
3.2.5.3 综合生命表
前述简单生命表虽有很多栏,但核心是lx栏(或nx栏,lx栏是标准化了的nx),其他各栏均可由它导出。综合生命表除lx栏,增加了mx栏,mx栏可称为生育力表(fecundity schedule),它描述了各年龄的出生率。综合生命表还增加了kx栏,kx栏表示各年龄组死亡率的指标,有的学者称为致死压力(killing power),它由lx导出,即kx=lglx一lglx+1。综合生命表同时包括了存活率和出生率两方面数据.将各年龄的lx与mx相乘,并累加起来,即可估计出一个非常有用的R0=∑(lxmx)值,称为净增殖率(net reproductive rate),同时R0还代表种群在生命表所包括特定时间中的世代净增殖率。在1年生生物中(没有重叠世代),R0表示种群在整个生命表时期中增长或下降的程度,R0>l,种群增长;R0=I,种群稳定;0<R0<l,种群下降。在表3-2中R0值为2.41,表明小天蓝绣球(Phlox drummondii)种群在增长。该生命表在种子发芽前进行种子数量统计以后,定期统计植物数量直至全部开花和死亡,其中各栏的符号和含义与前面表3-l藤壶生命表一样,只是x是以日为单位,ax栏与nx栏完全相当。
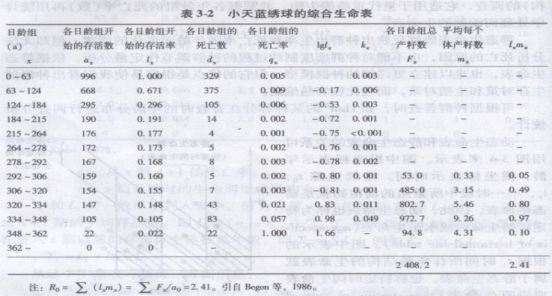
从mx栏看到,该植物有很长的繁殖前期,繁殖期开始生育力较低.以后逐渐升高,一旦到生育力高峰后就迅速下降,个体也接近死亡。lx栏说明前184天种群稀疏很快,中期死亡不多,开花后迅速死亡。正是出生率和存活数的联合变化,形成了Fx栏,它代表了各日龄组的总产籽数,所以∑Fx是该调查种群经一世代后的总产籽数。用此栏数字估计R0值与lxmx栏估计的是一样,R0=2.41表示经过一世代,小天蓝绣球的种子数平均增长到原来的2.41倍。
3.2.5.4 图解生命表
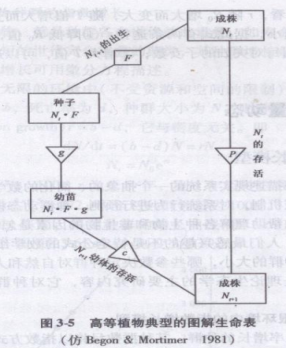
图解生命表(diagrammatic life table)是描述种群生死过程的另一种方式,尤其是对生活史比较复杂的种类。图解生命表最初用于高等植物,具有清晰、直观的特点。图3-5的3个方块表示1年生高等植物的种子、幼苗和成株3个时期,三角表示3个时期间的转化系数,如P为存活率、F为结籽率。
该图解生命表清晰表明生活史各阶段的生死过程。如t+1时间的成株Nt+1有明显两个来源:①前一时间成株数Nt的存活个体数,即Nt×P;②Nt的结籽数,经萌发并得以成长的个体数,即Nt×F×g×e。综合①、②得
![]()
不言而喻,这个模型假定没有迁入和迁出。这类图解生命表一般适用于世代不相重叠的1年生生物。
3.2.6 种群增长率r和内禀增长率rm
种群的实际增长率称为自然增长率(natural rate of increase),用r表示,不考虑种群的迁人和迁出,自然增长率可由出生率和死亡率相减计算出。世代的净增率R0虽是很有用的参数.但由于各种生物的平均世代时间并不相等,进行种间比较时R0的可比性并不强,两种群增长率(r)则显得更有应用价值。r可按下式计算:
r=lnR0/T
式中T为世代时间(generation time),是指种群中世代从母体出生到子代再产生的平均时间。用生命表资料可估计出世代时间的近时值,即
T=(∑xlxmx)/(∑lxmx)
自然界的环境条件在不断变化着,当环境条件有利时,r值可能是正值,条件不利时.可能变为负值。因此,长期观察某种群动态时,自然增长率r值是很有用的指标。但是为了进行比较,人们经常在实验室不受限制的条件下观察种群的内禀增长率(innate rate of increase),用rm表示。按Andrewartha的定义,rm是具有稳定年龄结构的种群,在食物不受限制,同种其他个体的密度维持在最适水平,环境中没有天敌,并在某特定的温度、湿度、光照和食物等的环境条件组配下,种群的最大瞬时增长率。因为实验条件下并不一定是“最理想的”,所以由实验测定的rm值不会是固定不变的。
从r=lnR0/T来看,r随R0增大而变大,随T值减小而变小。例如计划生育的目的是要使r变小,据此式有两条途径:降低R0值,即,使世代增长率降低,这就要求限制每对夫妇的子女数;增大T值,可以通过推迟首次生殖时间或晚婚来达到。
3.3种群的数量动态
3·3.1种群的增长模型
数学模型是用来描述现实系统的一个抽象的、简化的数学结构。人们用数学模型描述系统的内在机制,对系统行为进行预测。种群动态模型是自然种群动态和规律的简明描述,帮助理解各种生物和非生物的因素是怎样影响动态变化的。在数学模型研究中,人们最感兴趣的不是特定公式的数学细节.而是模型的结构,哪些因素决定种群的大小,哪些参数决定种群对自然和人为干扰的反应速度等。种群动态模型是理论生态学的主要研究内容,它对种群生态学做出了重要贡献。
3.3.1.1种群在无限环境中的指数增长模型
一个以内禀增长率增长的种群,其种群数目将以指数方式增加,只有在种群不受资源限制的环境下,这种现象才会发生。尽管种群数量增长很快,但种群增长率不变,不受种群自身密度变化的影响,这类指数生长称为与密度无关的种群增长(density-independent growth)或种群的无限增长。指数增长模型可分为两类:
(1)离散世代生物种群的指数增长
所谓离散世代生物,就是种群各个世代不相重叠,如许多1年生植物和昆虫,种群增长是不连续的,种群没有迁入和迁出,种群的增长为几何级数方程。即
Nt+1=R0Nt
式中 Nt--------叶世代种群大小;
Nt+1--------t+1世代种群大小;
R0----------世代净增率(周限增长率)。
如果种群以R0速率年复一年地增长,即
Nt=N0Rt0
将方程式Nt=N0Rt0两侧取对数,即
lgNt=lgN0+tlgR0
这是直线方程y=a+bx的形式,因此以lgNt对t作图,就能得到一条直线,其中lgN0是截距,lgR0是斜率。
R0是种群离散增长模型中的重要参数,R0>1,种群上升;R0<1,种群稳定;0<R0<1,种群下降;R0=0,雌体没有繁殖,种群在下一代灭亡。
(2)重叠世代生物种群的指数增长
所谓重叠世代生物,就是种群各个世代彼此重叠,如寿命很长的树木、人和多数兽类、生活史极短的世代完全重叠的菌类,其种群增长是连续的,种群没有迁人和迁出,种群的增长可用微分方程描述。
假定种群生活在无限的环境中(不受资源和空间的限制),在很短时间dt内种群的瞬时出生率为b,死亡率为d,种群大小为N,则种群的每员增长率(per-capita rate of population growth )r=b‐d,它与密度无关。即
dN/dt=(b一d)N=rN
其积分式: Nt=N0en
例如,初始种群N0=100,r为0.5/年,则1年后种群数量为100e0.5=165;2年后种群数量为100e2×0.5=272;3年后种群数量为100e3×0.5=448。
以种群Nt对时间t作图,得到种群的增长曲线[图3-6(b)],显然曲线呈J字型,但以lgNt对t作图[图3-6(a)],则变为直线。
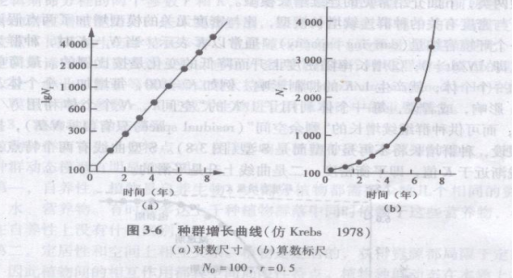
r是一种瞬时增长率,r>0,种群上升;r=0,种群稳定;r<0.种群下降。
3.3.1.2 种群在有限环境中的逻辑斯谛增长模型
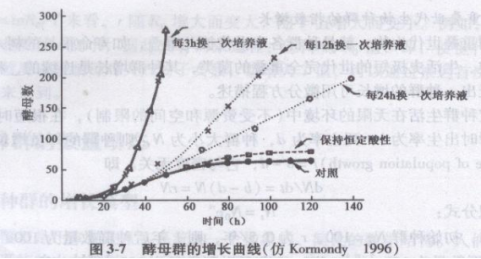
因为环境是有限的,生物本身也是有限的。所以大多数种群的J字型生长都是暂时的,一般仪发生在早期阶段,密度很低,资源丰富的情况下,而随着密度增大,资源缺乏。代谢产物积累等,由竞争、疾病、胁迫等引起的环境压力势必影响到种群的增长率r,使r降低。图3-7为不同方式培养酵母细胞时酵母实验种群的增长曲线,每3h换一次培养基,代表种群增长所所需营养物资源不受限制时的状况,显然此时的种群增长曲线为呈J型的指数增长。随着更换培养液的时间延长,种群增长逐渐受到资源限制,增长曲线也渐渐由J型变为S型,这就是下面介绍的种群在有限环境下的增长曲线。
受自身密度影响的种群增长称为与密度有关的种群增长(density-dependent growth)或种群的有限增长。种群的有限增长同样分为离散增长模型和连续增长模型两类,下面介绍常见的连续增长模型。
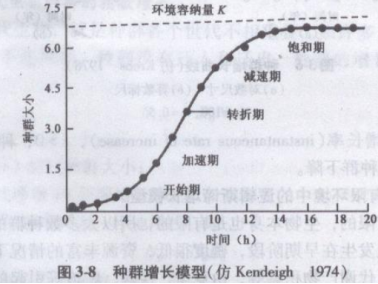
与密度有关的种群连续增长模型,比与密度无关的模型增加了两点假设:①有一个环境容纳量(carrying capacity),通常以K表示,当Nt=K时,种群为零增长,即dN/dt=0;②增长率随密度上升而降低的变化是按比例的。最简单是每增加一个个体,就产生l/K的抑制影响。例如K=100,每增加一个个体,产生0.01的影响,或者说,每一个体利用了l/K的“空间”,N个个体利用N/K“空间”,而可供种群继续增长的“剩余空间”(residual space)只有(1一N/K),按此两点假设,种群增长将不再是J型而是S型(图3-8)。S型曲线有两个特点:一是曲线渐近于K值,即平衡密度;二是曲线上升是平滑的。
产生S型曲线的最简单数学模型可以指数增长方程乘上一个密度制约因子(1一N/K),就得到生态学发展史上著名的逻辑斯谛方程(logisticequation,或称阻滞方程),是1838年由Verhurst提出的。即
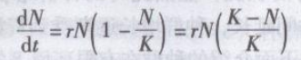
其积分式为
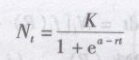
式中 a------参数,其值取决于N0,是表示曲线对原点的相对位置。
在种群增长早期阶段,种群大小N很小,N/K很小,因此,1一N/K接近于1,所以抑制效应可以忽略不计,种群增长实质为rN呈几何增长。然而.当N变大时,抑制效应增加,直到N=K时,(1一N/K)变成丁(1一K/K)=0,这时种群增长为零,种群达到了稳定的大小不变的平衡状态。逻辑斯谛曲线常划分5个时期:①开始期(initial phase),也可称为潜伏期(1atent phase)。此期间种群个体最少,密度增长缓慢,这是因为种群数量在开始增长时还很低。②加速期(accelerating phase),随个体数增加,密度增长逐渐加快。③转折期(inflecting phase),当个体数达到饱和密度一半(即K/2)时,密度增长最快。④减速期(decelerating phase),密度增长逐渐减慢。⑤饱和期(asymptotic phase),种群密度达到环境容纳量(K)而饱和。
逻辑斯谛方程的两个参数r和K,均有重要的生物学意义。如前所述,表示物种的潜在增殖能力,K是环境容纳量,即物种在特定环境中的平衡密度。但应该注意K同其他生态学特征一样,也是随环境条件(资源量)改变而变化。
逻辑斯谛方程的重要意义是:它是许多两个相互作用种群增长模型的基础:它是农业、林业、渔业等领域确立最大持续产量(maximum sustained yield)的主要模型;模型中两个参数r、K已成为生物进化对策理论中的重要概念。
3.3.1.3 植物种群动态模型
植物种群有下列4个特点,它直接影响了植物种群动态模型的建立,使其与动物种群动态模型有明显的不同。
第一,自养性。植物是自养生物.大部分植物都需要少数几个相同的资源.如光、水、营养物。有时候多达上千种植物群落中同时依赖于这些营养物,各种植物在自养性上没有什么区别。
第二,定居性和空间上相互关系。植物是定居的,获得资源都局限于定居地周围。因此植物间的相互作用都具空间局限性特点。植物种群动态在本质上与空间有密切关系,或者说,植物种群的平均增长率是空间分布的函数。同时,空间异质性对于植物种群有很大影响,甚至包括小尺度空间异质性对单个种子发芽的影响。
第三,生长的可塑性。前面已经提到,植株问生物量相差很大,因此其产籽数量可相差若干数量级。
第四,营养繁殖。许多植物,包括某些群落中的优势种有营养繁殖的能力。同一合子发育的个体(genet)上的许多构件(ramet)之间彼此有竞争资源的现象。
植物种群动态的模型研究,尚处于发展的早期。下面仅介绍2个模型:
(1)Tilman模型
Tilman模型是一种捕食者—被捕食者模型的扩展。Tilman把植物的密度视为捕食者的密度,而非生物资源视为被食者,从而建立了下面模型:
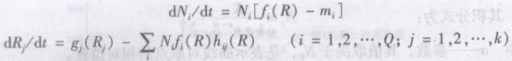
式中Ni——第i种密度;
fi(R)——出生率,死亡率
Rj——第j种资源;
gj(Rj)——该种资源在没有植物存在似的填补量或减少量;
hij(R)——形成i植物个体所需要的资源量。
(2)Skellam模型
这是考虑植物个体空间位置和个体间空间相互作用的模型。模型把生境划分为许多独立的小室,各小室内只具有一个成熟的个体,但加入了各小室之间的资源竞争,其模型为:
![]()
式中Xii——第i种在t时占有小室的比例;
di——i种的死亡率;
Pij——i种占有由于j种死亡而变空的小室的概率。
3.3.2自然种群的数量动态
野外种群不可能长期地连续地增长。要掌握种群动态规律,必须有长期的种群数量变动记录。自然界生物种繁多,而有长期种群动态记录的不多,主要是一砦具重大经济价值物种。研究种群动态的目的,是人类更好地利用和保护生物资源。
一种生物进入和占领新栖息地,首先经过种群增长和建立种群,以后可出现不规则的或规则的(即周期性)波动,也可能较长期地保持相对稳定;许多种类有时会出现骤然的数量猛增,种群大暴发,随后又是大崩溃;有时种群数量会出现长时期的下降(称为衰落),甚至死亡。
3.3.2.1 种群增长
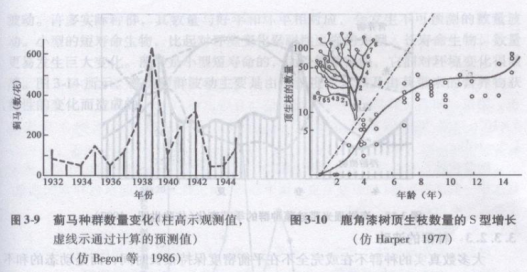
自然种群数量变动中,J型S型增长均可以见到,但曲线不像数学模型中所预测的光滑、典型.常常表现为两类增长型之间的中间过渡型。例如,澳大利亚昆虫学家Andrewartha曾对生活在玫瑰上的蓟马(Thrips imaginis)种群进行长达14年的研究(图3-9)。他发现.在环境条件较好的年份其数量增加迅速,直到繁殖结束时增加突然停止,表现出J型增长,但在环境条件不好的年份则是S型增长。对比各年增长曲线,可以见到许多中间过渡型。因此,J型增长可以视为是一种不完全的J型增长,即环境限制作用是突然发生的,在此之前,种群增长不受限制。
高等植物一个单株的构件数,如鹿角漆树(Rhus typhna)不同年龄的顶生枝数量,其增长也是S型(图3—10)。
3.3.2.2季节消长
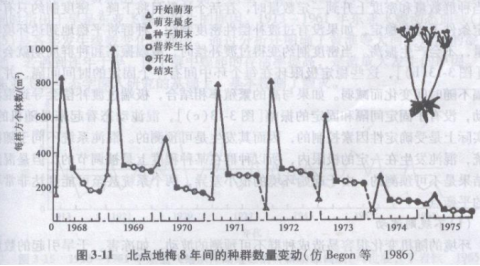
对自然种群的数量动态,首先应区别一年内(季节消长)和年间变动。1年生草本植物北点地梅(Androsace septentrionalis)种群个体数有明显的季节消长(图3-11)。8年间,籽苗数为500--l 000株/m2。,每年死亡30%一70%,但至少有50株以上存活到开花结实,产出次年的种子。因此,各年间的成株数量变动很少。
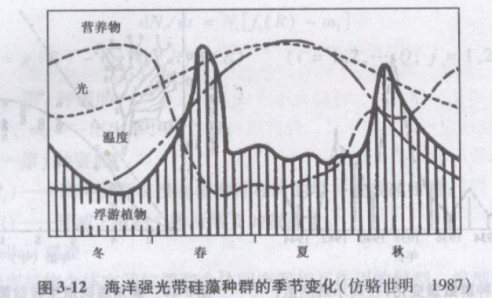
季节性消长,主要是环境季节性变化引起的,如海洋强光带的硅藻种群的季节变化(图3-12),往往每年有春秋两次密度高峰称为“开花”(bloom)。其原因是冬季的低温和光照减少.降低了水体的光台强度,营养物质随之逐渐积累;到春季水温升高、光照适宜.加之有充分营养物,使具巨大繁殖能力的硅藻迅速增长,形成春季数量高峰。但是不久以后营养物质耗尽,水温也可能过高,硅藻数量下降;以后营养物质又有积累,形成秋季较高的数量高峰。这种典型的季节消长,也会受到气候异常和人为的污染而有所改变。掌握其消长规律,是水体富营养化预测和防治所必须的。
3.3.2.3种群的波动
大多数真实的种群不在或完全不在平衡密度保持很长时间,而是动态的和不断变化的。因为以下几个原因,种群可能在环境容纳量附近波动:①自然界环境的随机变化。因为随着环境条件如天气的变化,环境容纳量就会相应的变化。②时滞或称为延缓的密度制约。在密度变化和密度对出生率和死亡率影响之间导入一个时滞,在理论种群中很容易产生波动,种群可以超过环境容纳量.然后表现出缓慢的减幅振荡直到稳定在平衡密度[图3-13(a)]。③过渡补偿性密度制约,即当种群数量和密度上升到一定数量时,存活个体数目将下降。密度制约只有在一定条件下才会稳定.如果没有过渡补偿性密度制约,种群将平稳地到达环境容
纳量,不会产生振荡。当密度制约变得过渡补偿时,减幅振荡和种群周期就会发生[图3-13(b)],这些稳定极限环在每个环中间有一个固定的时间间隔,并且振幅不随时间变化而减弱。如果与高的繁殖率相结合,极端过渡补偿会导致混沌波动,没有了固定间隔和固定的振幅[图3-13(c)],混沌动态看起来是随机的,但实际上是受确定性因素控制的,因而其发生是可预测的。混沌系统不同于随机系统,混沌发生在一定的极限内,所以种群在某种程度上是被调节的,但是混沌的结果是不可预测的。由于起始环境的很小差异,两个系统甚至可能到达非常不同的平衡点。
(1)不规则波动
环境的随机变化很容易造成种群不可预测的波动,如冻害、干旱引起的数量波动。许多
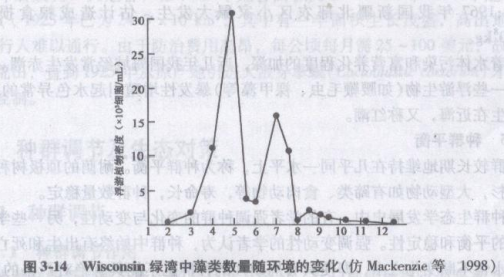
实际种群,其数量与好年和坏年相对应,会发生不可预测的数量波动。小型的短寿命生物,比起对环境变化忍耐性更强的大型、长寿命生物,数量更易发生巨大变化。藻类是小型短寿命的,而且繁殖很快,它们对环境变化很敏感。图3-14所示,藻类种群波动主要是由于温度变化以及由其带来的营养物获得性的变化而造成的。
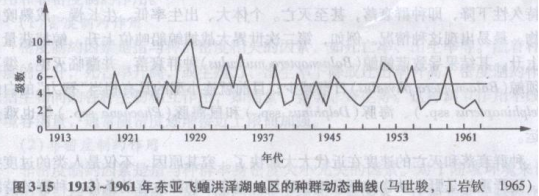
马世骏(1985)探讨过大约1000年有关东亚飞蝗危害和气象资料的关系,明确了东亚飞蝗在我国的大发生没有周期性现象,同时指出于旱是东亚飞蝗大发生的原因。1913~1961年东亚飞蝗动态曲线(图3-15)是各年发生级数变化序列图。通过分析还明确了黄淮等大河三角洲的湿生草地,若遇到连年干旱.使土壤中蝗卵的存活率提高,是造成大发生的原因。在对东亚飞蝗生态学深入研究的基础上,基本控制了蝗灾。
(2)周期性波动
种群的周期性波动,包括季节性波动和年波动。在一些情况下,捕食与食草作用等导致延缓的密度制约会造成种群的周期性波动。经典的例子为旅鼠、北极狐的3~4年周期和美洲兔、加拿大猞猁的9~10年周期。几乎每一本动物生态学书都有关于这两类周期性波动的描述,生态学对周期性数量波动的讨论也特别热烈。根据近30年资料,我国黑龙江伊春林区的小型鼠类种群也有明显3~4年周期,主要是优势种棕背鼠平(Cletnrionomys rufocanus)每遇高峰年的冬季就造成林木危害,尤其是幼树,对森林更新危害很大;并且,其周期与红松结实的周期性丰收相一致。根据以鼠为主要食物的黄鼬(Mustela sibirica)的每年毛皮收购记录,证明黄鼬也有3年周期性,但高峰比鼠晚一年。
3.3.2.4 种群暴发
具不规则或周期性波动的生物都可能出现种群的暴发。最著名的暴发见于害虫、害鼠。如索马里1957年一次蝗灾估计有蝗虫约160×l08只之多。总重量达5×104;1967年我国新疆北部农区小家鼠大发生,估计造成粮食损失达1. 5×108kg。
随着水体污染和富营养化程度的加深,近几年我国海域经常发生赤潮。赤潮是水中一些浮游生物(如腰鞭毛虫、裸甲藻等)暴发性增殖引起水色异常的现象,主要发牛在近海.又称红潮。
3.3.2.5 种群平衡
种群较长期地维持在几乎同一水平上,称为种群平衡。耐荫的顶极树种如云杉、冷杉,大型动物如有蹄类、食肉动物等,寿命长,种群数量稳定。
在种群生态学发展史中,有的学者强调种群的变化与变动性,另一些学者强调种群的平衡和稳定性。强调变动性的学者认为,种群中始终有出生和死亡,总是处在永恒不断变化之中,即使具有周期性的种群也不能具有绝对严格的周期。强调稳定性的学者则认为,每个种群一般均有一平均密度和平衡水平,当种群数量偏离此水平时,种群就有重新返回原有水平的倾向,即使具有强烈数量波动特点的种类,如旅鼠和北极狐等,其种群的周期性(即几乎定期的升降)也可看作是一种稳定性。
3.3.2.6 种群的衰落和死亡
当种群长久处于不利条件下(人类过伐、过捕或栖息地破坏),其数量会出现持久性下降,即种群衰落,甚至灭亡。个体大、出生率低、生长慢、成熟晚的生物,最易出现这种情况。例如,第二次世界大战捕鲸船吨位上升,鲸捕获量节节上升,其结果导致蓝鰮鲸种群衰落,并濒临灭绝;继而长须鲸日渐减少;目前就连小型的具有相当“智力”的白鲸、海豚和鼠海豚等也难逃厄运。
种群衰落和灭亡的速度在近代大大加快了。究其原因,不仅是人类的过度捕杀,更严重的是野生动物的栖息地被破坏.剥夺了物种生存的条件。种群的持续生存,不仅需要保护良好的栖息环境,而且要有足够的数量达到最低种群密度。因为过低的数量会因近亲繁殖而使种群的生育力和生活力衰退。保护生物学研究的一个热点问题,就是进行下降种群的生存力分析,判断最小可存活种群(minimum viable population),即种群以一定概率存活一定时间的最小种群的大小。
3.3.2.7 生态入侵
由于人类有意识或无意识地把某种生物带人适宜其栖息和繁衍的地区,该生物种群不断扩大,分布区逐步稳定地扩展,这种过程称为生态入侵(ecological invasion)。如紫茎泽兰(Eupatorium adenophorum)原产墨西哥,新中国成立前由缅甸、越南进入我国云南,现已蔓延到25°33’N地区,并向东扩展到广西、贵州境内,它常常连接成片,发展成单种优势群落,侵入农田,危害牲畜,影响林木生长,成为当地“害草”。仙人掌原产美洲,有数百种,其中26种被引人澳大利亚作为园艺植物,于1839年引进做篱笆的仙人掌因扩展迅速,1880年后成为一大危害,在1890年危害面积为4×104km2,1920年达23.5×104km2,1925年已为24.3×104km2,其中有一半面积生长茂盛,高出地面l~2m,行人难以通行。由于防治费用高昂,每公顷每月需25~100美元,故多被废弃为荒田,直到1925年从原产地引进天敌穿掌螟才使危害得以控制。
3.4种群调节及生态对策
3.4.1种群调节
3.4.1.1 种群调节作用
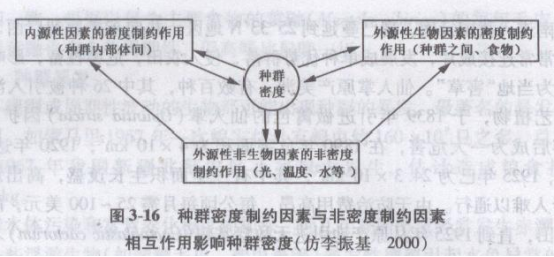
当种群数量偏离平衡水平上升或下降时,有一种使种群返回平衡水平的作用,称为种群调节。种群调节使种群具有一定的稳定性,能够减少波动,保持在一个稳定的数量上。在自然界中,种群密度的极端值是很少达到的,因为有一系列的机制限制种群的增长。种群调节以种群密度为基础,但有时种群数量的变动与密度无关,而是受外界因素的影响,所以通常把影响种群调节的各种因素分为两大类,即密度制约和非密度制约因素.而对种群的调节作用分别称为密度制约作用和非密度制约作用。
(1)密度制约作用
密度制约因素是指与种群密度相关的因素,如死亡率、出生率等。随着种群密度的上升,死亡率升高,或生殖力、出生率下降或迁出率升高。密度制约作用包括生物间的各种生物相互作用,如捕食、竞争、寄生等。这种调节作用不改变环境容纳量,通常随密度逐渐接近上限而加强。
(2)非密度制约作用
非密度制约因素是指与种群本身密度大小无关的因素。对于陆域环境来说,这些因素包括光照、温度、风、降雨等非生物的气候因素。这种非密度制约作用是通过环境的变动而影响环境容纳量,从而达到调节作用。
在实际研究中,这两类因素往往相互联系,难以分开。例如气候影响食物供给,食物影响出生率,从而影响种群密度,种群密度决定于生物与环境关系的各个方面.并由此影响生物的密度(图3-16)。
3.4.1.2 种群调节理论
种群的这种调节是通过种群本身个反向力之间的平衡而达到的。限制这两的内禀增长率,它在不受限制的条件下,对任何种群都可导致指数增长,另一方面种群的大小受气候、
 资源、竞争者、捕食者、寄生者或疾病等环境因素所决定,以上两方面的作用,表现为出生率和死亡率的平衡。因此,影响出生率和死亡率、迁人和迁出的一切因素.都同时影响种群的数量动态变化,于是生态学家提出了许多不同的假说来解释种群的动态机制。
资源、竞争者、捕食者、寄生者或疾病等环境因素所决定,以上两方面的作用,表现为出生率和死亡率的平衡。因此,影响出生率和死亡率、迁人和迁出的一切因素.都同时影响种群的数量动态变化,于是生态学家提出了许多不同的假说来解释种群的动态机制。
(1)气候学论
气候学论强调非密度制约因索,提出气候是调节昆虫种群密度的因素。如以色列的F.S. Bodenheimer(1928)认为,天气条件通过影响昆虫的发育和存活决定种群密度。证明昆虫早期死亡率的85%~90%是由于气候条件不良而引起的。气候学论多以昆虫为研究对象,认为生物种群主要是受对种群增长不利的短暂气候所限制。因此.从来就没有足够的时间增殖到环境容纳量所允许的数量水平,不会产生食物竞争。
(2)生物学论
生物学论主张捕食、竞争、寄生等生物过程对种群调节起决定作用。如澳大利亚生物学家A.J.Nicholson虽然承认非密度制约因子对种群动态有作用,但认为这些因子仅仅是破坏性的,而不是调节性的。他举例说明:假设一昆虫种群世代增加100倍,而气候变化消灭了98%.那么该种群仍然要每世代增加l倍,但如果存在一种昆虫的寄生虫,其作用随昆虫密度的变化而消灭了另外的1%,这样种群数量得以调节并能保持稳定。在这种情况下,寄生造成的死亡率虽小却是种群的调节因子。
Smith支持nicholson的观点,指出种群呈现出两个主要的种群增长现象:一是随时间变化的趋势.另一个是这一变化围绕一个平均密度而变化的趋势。这一平均密度是某一特定环境的特征,不同的环境其平均密度不同。Smith认为,在决定种群平均密度的诸因子中,寄生物、捕食动物,以及疾病起了最为重要的作用。
强调食物因素对种群的调节也可以归入生物学论。例如。英国的鸟类学家D.Lack(1954)认为,种群调节原因可能有3个:食物的短缺、捕食和疾病。其中食物是决定性的。Pitelk.(1964)与Schultz(1964)提出营养恢复假说(nutrient recovery hypothesis),说明在阿拉斯加荒漠上,旅鼠(Lemmus trimucronatus)的周期性变化是植食动物与植被间相互作用所导致的,在旅鼠数量很高的年份,食物资源被大量消耗,植被量减少,食物的质(特别是含磷量)和量下降,幼鼠因营养条件恶化而大量死亡以至种群数量下降,低种群密度使植被的质和量逐步恢复,种群数量再回升,周期大约3~4年。
(3)协调派的折衷观点
20世纪50年代气候学派和生物学派发生激烈论战,但有的学者提出了折衷的观点。如A. Milne即承认密度制约因子对种群调节的决定作用,也承认非密度制约因子具有决定作用。他把种群数量动态分成了3个区:极高数量、普通数量和极低数量。在对物种最有利的条件下,种群数量最高,密度制约因子决定种群数量;在环境条件恶劣的条件下,非密度制约因子左右种群数量变动。折衷观点认为,气候学派和生物学派的争论反映了他们工作地区环境条件的不同。
(4)自动调节学说
上述学派的研究焦点都集中在外源性因子,主张自动调节的学者则将研究焦点放在动物种群内部,即内源性因子,其特点包括强调种内成员的异质性,异质性可能表现在行为上、生理上和遗传性质上;认为种群密度的变化影响了种内成员,使出生率、死亡率等种群参数变化;主张把种群调节看成是物种的适应性反应,它经自然选择,带来进化上的利益。
3.4.1.3 林分密度调节
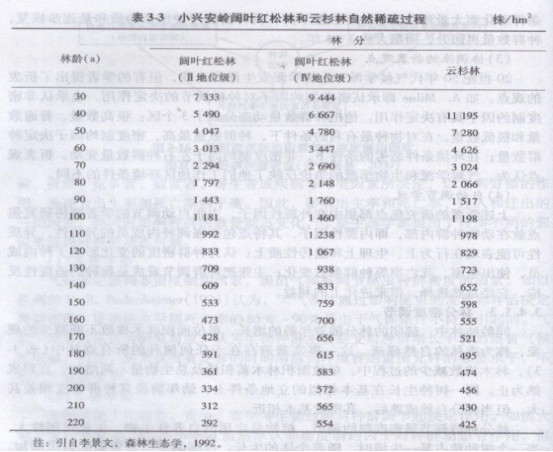
同龄纯林中,郁闭的林分随着年龄的增长,单位面积林术株数不断减少的现象,称为森林的自然稀疏。这一事实普遍存在于任何树种的所有森林中(表3-3).林木株数减少的过程中,单位面积林木蓄积量及总生物量不断增加,直到成熟为止。同一树种生长在基本相似的立地条件上,幼年阶段其种群密度相差甚大,但当林分自然成熟后,其密度基本相近。
林分种群调节属密度制约调节,植物是定居的自养性生物,生长可塑性大,当一个树种独占某一生境时,随着个体的生长,占据的空间和资源的需求增加。这时林分种群面临两种选择:一是降低每株林木获得水分、养分及光照的能力,以每株林木的缓慢生长,换取现有种群数量(密度)不变。林木缓慢生长的结果是抵抗自然灾害的能力下降,在生态环境大幅度波动下(如干旱),种群可能崩溃。二是任何生态系统自然进化的方向是增加稳定性,因此选择不断减少林木株数来保证林分稳定性,把有限的资源集中到优良个体上。林木个体的遗传性存在差异,这种差异在种子阶段就存在,大粒饱满的种子往往形成强壮的幼苗,林地土壤非均匀性及微生境的异质性,与林木个体遗传性结合,为林术分化提供了基本条件。因而,生长弱的个体不仅得不到足够营养物质满足生长发育的需要.而且环境空间也愈来愈小,适应能力和竞争能力的减弱,使它们逐渐死亡而被淘汰,使所保留的林木扩大生长空间,这个过程从林分郁闭起一直到自然成熟始终连续进行。
林分密度调节的核心是自然稀疏,即不断减少林木株数调节生长和繁殖。林木密度调节是一种进化适应,当立地条件对林木生长极端不利时,林木生长受到物理环境的强烈抑制,林分始终不郁闭,个体林木间没有竞争,这时表现不出密度调节。如大兴安岭的苔藓落叶松
林。当永冻层距地面很近时,由于土壤低温,林木生长十分缓慢,林术枝叶稀疏。影响林木死亡的因素是外因作用,与林分密度无关,非密度制约因素起主要作用。树种生长在较好的生境上才出现密度调节,这时林分充分郁闭,发生空间和资源的竞争,密度调节才发挥作用。林分密度越大,营养物质供应越紧张,林木间的竞争越激烈,密度调节表现越明显,单位时间死亡的林木株数就越多。相同密度的同龄纯林在不同立地条件上,密度调节发生的时间和强度有差异。良好的立地条件,林木生长迅速,旺盛生长的林术需要更多的营养物质和空间,密度调节生长的时间较早,调节高峰出现得早,单位时间死亡的林木株数多。喜光树种形成的林分,自然稀疏早、强度大,调节高峰出现得早。
林木生长对密度有较大可塑性反应,孤立木有庞大的树冠,树高与直径比值小,树干尖削度大;密林中林木树冠狭小,树高与直径比值较大,干形良好。同理,密林中单株林木结实量少,种子质量欠佳,生长的可塑性反应还表现在下一个世代的更新与生长,密林下更新的幼苗幼树生长缓慢,在长达46年中没有直径生长。连续自然稀疏的结果使林分成熟时,单位面积林术株数减少。虽然密度有较大的变幅,单位面积上种子产量仍保持相对稳定,林木通过外部动态变化进行调节,如红松林,成熟前有明显的顶端优势。但成熟时,中央顶枝生长缓慢,侧枝加速生长,出现所谓“平头”,增大树冠体积、产生更多球果,弥补成熟时单位面积株数少,可能导致的球果产量下降。
另一种情况,暂时观察不到自然稀疏的发生,小面积上,当乔木树种幼中龄个体在地面上的分布非常均匀,土壤微生境条件差异也很小,或者接近相等时,所有个体都受到强烈的抑制,林木个体生长潜力差异不能表现出来,激烈分化和自然稀疏在短期内观察不到。但这种现象不能长久维持。可能有两种结果:一是出现外伤性干扰或有害因子作用时全部毁灭.另外可能终将出现分化和稀疏。
林分的密度效应在林业生产中具有极大的经济效益和生态效益,人们把注意力集中在适宜密度上来.以期找出树种的各年龄阶段的适宜密度,定量地表示密度与产量关系,用以指导农、林业生产。
3.4.2 生态对策
生物的生活史(1ife history)就是指从出生至死亡所经历的全部过程。生活史的关键组分是身体大小(body size)、生长率(growth rate)、繁殖(reproduction)和寿命(longevity)。生物在其漫长的进化中,分化出各种各样的生物有机体。不同种类的生物生活史类型存在巨大变异。一些种类能活至成百上千年,如红豆杉(Taxus baccata),一些个体巨大如大象、鲸和加利福尼亚红杉(Sequoia sempervirens),一些个体寿命短且身体微小如真菌、大肠杆菌等。生物在生存斗争中而朝不同方向进化获得的生存对策称生态对策(bionomic strategy)或生活史对策(life history strategy)。例如,生殖对策、取食对策、逃避捕食对策、扩散对策等。而r-对策和K-对策关系到生活史整体的各个方面。
3.4.2.1 r-对策和K-对策
R.H.MacArthur(1962)总结了前人对生物生活史的研究,认为热带雨林的气候条件稳定,自然灾害较为罕见,动物的繁衍有可能接近环境容纳量,即近似于逻辑斯谛方程中的饱和密度(K)。故在稳定的环境中,谁能更好地利用环境承载力,达到更高的K值,对谁就有利。相反,在环境不稳定的地方和自然灾害频繁发生,只有较高的繁殖能力才能补偿灾害所造成的损失。故在不稳定的环境中.谁具有较高的繁殖能力将对谁有利。通常用来表达繁殖力的测度之一是内禀增长率rm,所以居住在不稳定环境中的物种,具有较大的rm是有利的。有利于增大内禀增长率的选择称为r-对策,有利于竞争能力增加的选择称为K-对策。R .H . MacArthur和E . O . wilson(1967)又从物种适应性,进一步把r-对策的物种称为r-策略者(r-strategistis),K-对策的物种称为K-策略者(K-strategistis),他们认为,物种总是面临两个相互对立的进化途径,各自只能选择其一,才能在竞争中生存下来。美国生态学家E .R .Pianka (1970)将r-对策和K-对策理论推广到一切有机体,并将两种选择的特征总结于表3-4。

r-策略者是新生境的开拓者,但存活要靠机会,所以在一定意义上,它们是机会主义者,很容易出现“突然的暴发和迅速的破产”。r-策略者个体小,寿命短,对后代的“投资”不注重其质量,更多是考虑数量。在植物中,1年生植物和农田杂草、原生和次生裸地的先锋种属于r-策略者。而K-策略者是稳定环境的维护者,在一定意义上,它们又是保守主义者,当生存环境发生灾变时很难迅速恢复,如果再有竞争者抑制,就可能趋向灭绝。K-策略者个体太,寿命长,高的竞争能力以及对后代的巨大“投资”,因为它们硕大的体型对环境灾变缺乏相应的适应能力。因此K-策略者要良好的生长,必须在稳定的生境条件下,进化方向是在稳定条件下增强种问竞争能力,选择大型个体是有利的,但种的扩散能力低,进化压力使种群保持或接近K值,种群增长率r较小,而保持高的存活率,必须在防御机制上给予很大“投资”,占有较大比例能量,用于生长繁殖的能量少,但利用能量的效率高,如秃鹰能做长距离的滑翔。大多数森林树种属于K-策略者。
自r-K对策理论和概念提出以来,有关学者开展了广泛的研究。在实际应用中,这一理论既用于较大类群之间的比较,也用于近似物群之间的比较,甚至于同一物种之内不同类型和不同生境个体之间的比较。例如,森林树木和大型哺乳动物具有K-对策特征,而1年生植物和昆虫一般具有r-对策特征。支持r/K二分化的一个很好的例子来自两种香蒲属植物分别在美国德克萨斯州和北达科他州,北达科他州的T . angustifolia与德克萨斯州的T .domingensis相比经历冬季高死亡率和低竞争,如r/K学说预测的那样,T . angustifolia较T . domingensis成熟更早(44天/70天)、体形较矮(162cm/186cm).并且结果更多(每株41个/8个)。同一植物种群的不同个体,在不同立地条件上的生长.亦有不同生态对策趋向。在研究蒲公英生活史时,发现3个种群中可识别出4个独立的无性系,分别称为A、B、C、D,A、B、C在3个种群中都出现,D未出现在干扰最大的立地上,研究者认为A是典型的r-对策种,在严重干扰的立地(小路)上最丰富,在稳定的立地(老牧场)上最少,A在单一栽培时有较高的繁殖产量,D是典型的K-对策种,它在干扰最小种群中最丰富,B和C处在中间状态。从极端的r-策略者到极端K-策略者之间,中间有很多过渡的类型,有的更接近r-对策,有的更接近K-对策,这是一个连续的谱系,可称为r-K连续体(r-K continuum of strategies )。
另外,同一物种分布在不同生态梯度上也可形成一种r-K连续特征。例如,云杉在低海拔属于偏K-对策,中海拔为K-对策,中高海拔为偏K-对策,高海拔为r-对策(江洪 1992)。
但是,也有许多事例不支持r/K二分法,例如,在所有大小相似的动物中,蚜虫具有较高的种群增长率(表明它们是r-策略者)却生育较大型的后代(一个K-对策特征)。现在一般不认为r/K置理论是错误的,而认为这是一种特殊情况,被具有更广预测能力的更好的模型所包含。
r-对策和K-对策在进化中各有其优缺点。K-对策种群竞争性强,数量较稳定,一般稳定在K附近,大量死亡或导致生境退化的可能性较小,但一旦受危害造成种群数量剧烈下降,在动物保护中应特别注意。相反,r-策略者死亡率甚高,但高r值使种群能迅速恢复。而高的扩散能力还可使其迅速离开恶化环境,在其他地方建立新的种群。r-策略者的高死亡率、高运动性和连续地面临新环境,更有利于形成新物种。
3.4.2.2 R -、C - 和S-对策的生活史式样
20世纪70年代后期,英国生态学家J .P . Grime等在r-对策和K-对策的基础上对生活史式样的分类作了有益的补充。除上面提到的r-对策和K-对策概念外,还提出了多种划分生境的方案,以试图建立一种连接生境与生活史的模式。这些分类方案必须能够划分所有生境,并且要从正在讨论的生物的角度出发来分类。因为一个生境是均质性还是异质性的。是良好还是恶劣的,会因所讨论的生物的不同而不同。
例如,可将生境划分为导致高繁殖付出(高-CR)的生境和导致低繁殖付出(低-CR)的生境。在高-CR生境(那里竞争剧烈,或对小型成体捕食严重),任何由于繁殖而导致的生长下降都会使未来繁殖付出高代价。因此可预期,在高-CR生境中生活的物种,其繁殖会在达到一个适度的身体大小以后才开始。与此相反,在低-CR生境(此处竞争弱,大型个体处在较强的捕食压力下,或死亡率很高而且是随机的),推迟繁殖没有任何优势。
“两面下注”(“bet-hedging”)理论根据对生活史不同组分(出生率、幼体死亡率、成体死亡率等)的影响比较不同生境。如果成体死亡率与幼体死亡率相比相对稳定,可预期成体会“保卫其赌注”,在很长一段时期内生产后代(也就是多次生殖),而如果幼体死亡率低于成体,则其分配给繁殖的能量就应该高,后代一次全部产出(单次生殖)。
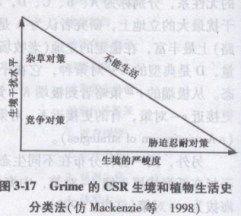
Grime的CSR三角形是对植物生活史的三途径划分.这比r/K二分法应用更广些。这种划分有两个轴,一轴代表生境干扰(或稳定性),另一轴代表生境对植物的平均严峻度。植物的潜在生境有3种类型:①低严峻度、低干扰;②低严峻度、高干扰;③高严峻度、低干扰。需要明确的是,生物在高严峻度、高干扰生境,如活跃的火山和高移动性的沙丘,是不能生活的。这3种生境的每一种都支持特定的生活史对策(图3-17)。低严峻度、低干扰生境支持成体间竞争能力最大化的生活史对策(C-对策);低严峻度、高干扰生境支持高繁殖率,这是杂草种类特有的杂草对策(R-对策);高严峻度、低干扰生境,如沙漠,支持胁迫忍耐对策(S-对策)。
第4章 林木种内与种间关系
【本章提要】生物在自然界长期发育与进化的过程中,表现了以食物、资源和空间关系为主的种内与种问关系,它们主要以竞争、捕食、寄生、共生而发生相互作用,种间不断协同进化,以适应改变了的外部环境条件。了解它们关系之间的复杂性,有利于培养学生对生物群落的创建、控制、利用和管理能力。
森林是由植物、动物和微生物构成的一个生物群落。不同生物共同生括在同一环境里,彼此必然发生着相互作用。林木种群内个体间的相互关系称为种内关系(intraspecific relationship),而将生活于同一生境中的所有不同物种之间的关系称为种间关系(interspecific relationship)。
种内个体间或物种间的相互作用可根据相互作用的机制和影响分类。主要的种内相互作用是竞争(competition)、自相残杀(cannibalism)和利它主义(altruism)等,而主要的种间相互作用是竞争、捕食(predation)、寄生(parasitism)和互利共生(mutualism)(表4-1)。

通常,种群的相互关系比较复杂,其复杂性主要受下列因素影响:种群的密度和个体生长发育状况;种群内部和种群之间个体和个体的直接影响;群落内部的环境变化,如地上部分小气候环境和地下部分土壤环境的变化;以及植物本身的生态幅、地理分布、生活型、竞争能力,植物分泌物,隐蔽作用,寄生作用等,所有这些都会直接、间接地影响种群内部和种群之间个体的相互关系。
从理论上讲,林木的种内、种间存在多种多样关系。但是,种之间发生的相互作用对一方没有影响,而对另一方有益如偏利共生(commensuailsm),或有害如偏害共生(amensualism)。以相互作用的影响是正(+)、负(一),还是中性(0)为基础划分相互作用可能会更方便(表4-2)。
 4.1竞争
4.1竞争
竞争是指在同种(种内竞争)或异种(种间竞争)的两个或更多个体间,由于它们的需求或多或少地超过了当时空间或共同资源供应状况,从而发生对于环境贽源和空间的争夺,而产生的一种生存竞争现象。
4.1.1种内竞争
同种个体间发生的竞争称种内竞争(intraspecific competition)。由于同种个体通常分享共同资源,种内竞争可能会很激烈。但种内资源需求可能存在年龄差异。它们对资源利用的普遍重叠程度,意味着种内竞争是生态学的一种主要影响力。从个体看,种内竞争可能是有害的,但对整个种群而言,因淘汰了较弱的个体,保存了较强的个体,种内竞争可能有利于种群的进化与繁荣。
物种内个体之间的竞争极其普遍地存在于自然界中。种内竞争明显受密度制约,在有限的生境中,种群的数量越多,对资源的竞争就越激烈,对每个个体的影响也越大,死亡率可能会升高而出生率下降。但是。在某些情况下,特别是在种群密度很低时,出生率可能会随密度而增长,而死亡率会下降。由于物种内竞争和密度是紧密相连的,即无论何时产生竞争,它都既来源于密度又作用于密度。因此,种内竞争具有调节种群数量的动态趋势。
4.1.1.1 密度效应
植物种群内个体间的竞争,主要表现为个体间的密度效应(density effect),反映在个体产量和死亡率上。在一定时间内,当种群的个体数目增加时,就必定会出现邻接个体之间的相互影响,称为密度效应或邻接效应(the effect of neighbors)。植物生长的可塑性大,这种可塑性一方面表现在个体的生长对外部非生物环境的响应,另一方面表现在种群内个体之间存在密切关系。如在植物稀疏、环境条件良好的情况下,枝叶茂密.构件数很多;相反在植株密生和环境不良的情况下,可能只有少数枝叶,构件数很少。
表示密度效应的方法主要有以下2种。
(1)最后产量衡值法则
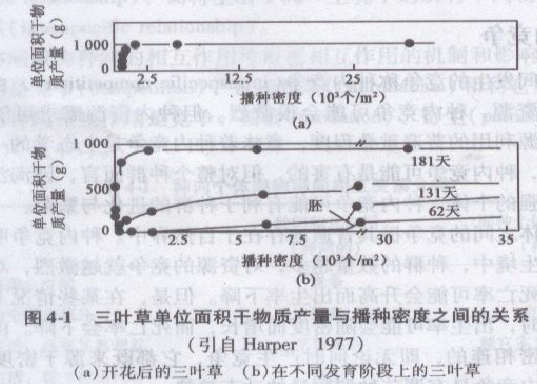
在一定范围内,当条件相同时,不管初始播种密度如何,最后产量差不多都是一样的,即最后产量衡值法则(law of constant final yield)。澳大利亚生态学家C . M . Donald(1951)对三叶草(Trifolium subterraneum)的密度与产量作了一系列研究后,证实了这个法则。图4-1表示单位面积上三叶草的干物质产量与播种密度的关系。其中,由图4-1(a)可以看出在密度很低时干物质随播种密度增加,但很快就趋于稳定;由图4-1(b)可以看出从萌芽初期到181天,都呈现出产量随密度的恒定的规律。
最后产量衡值法则可用下式表示
![]()
式中 ![]() ──植物个体平均重量;
──植物个体平均重量;
d——密度;
Y——单位面积产量;
Ki——常数。
最后产量衡值法则形成的原因是在高密度情况下,植株之间对光、水、营养物等资源的竞争十分激烈;在有限的资源中,植株的生长率较低,个体变小。
(2) -3/2自疏法则
随着播种密度的提高,种内竞争不仅影响到植株生长发育的速度,也影响到植株的存活率。在高密度的样方中。有些植株死亡了,这一过程称为自疏(self-thinning)。日本学者Yoda等(1963)把自疏过程中存学个体的平均株干重(![]() )与种群密度(d)之间的关系用下式来表示:
)与种群密度(d)之间的关系用下式来表示:
![]() =Cd-a
=Cd-a
两边取对数: 1g![]() =lgC – a lgd
=lgC – a lgd
英国生态学家J . L .Harper(1981)等对黑麦草(Lolium perenne)的研究发现,a为一个恒值等于3/2,因此![]() =Cd-3/2被称为 - 3/2自疏法则。White等(1980)对80多种植物的自疏作用进行测定,包括藓类、草本和木本植物等,都表现为-3/2自疏现象。
=Cd-3/2被称为 - 3/2自疏法则。White等(1980)对80多种植物的自疏作用进行测定,包括藓类、草本和木本植物等,都表现为-3/2自疏现象。
此关系式还适用于林分中的林木平均单株树干材积(V)与最大密度(dm)之间的关系:
V=Kdm-a
式中K,a——常数。
但通常a值变动不大,在3/2=l.5上下变动,这条线叫最大密度线,表示单株材积能够长成最大时的最大密度。日本几个树种平均单株干材积(V)的虽大密度(dm)线如下:
柳杉 V=l. 527×106dm-1.94
扁柏 V=l .517×105dm-1.74
落叶松 V=l. 046×104dm-1.50
常数a表示直线的斜率,此值与树种耐荫性有关,喜光树种较耐荫树种更接近3/2。
4.1.1.2 生态型
种群内不同个体属性在形态、生理、行为和生态方面几乎是一样的,因而竞争是最激烈的。自然选择的压力就迫使种群内个体发生变异、分化,扩大其分布范围。
同物种植物(树种)的不同个体群分布和生长在不同的生境里,由于长期受到不同生境条件的影响,在植物的生态适应进化过程中,就发生了不同个体群之间的变异和分化,形成了一些在生态学上互有差异的、异地性的个体群。这些差异在自然选择的过程中通过遗传性被固定下来,形成了在生态特性上有差异的不同个体群类型称为生态型(ecotype)。生态型的名词和概念是瑞典学者Turesson(1922)提出的:“一个种对某一特定生境发生基因型反应的产物”。简言之,生态型就是同种植物中生态学特性具有某些差异的类型。一般地说,生态分布区域广的种类生态型也多,具有更多生态型的种就能够更好地适应于广阔范围的生态环境变化。对树种的生态型研究直接关系到树种的选择、育种和引种工作,具有很重要的生产实践意义。
生态型主要有以下3大类型。
(1)气候生态型
当树种分布区扩展或栽种到不同气候地区,主要由于长期受气候因子的影响所形成的生态型。例如,在我国亚热带分布很广的马尾松可分为4个类型:北亚热带地理类型;中亚热带地理类型;南亚热带地理类型;四川盆地丘陵地理类型。
不同的气候生态型在形态、生理、生化上都表现有差异,如对光周期、温周期和低温春化等却有不同的反应。分布在南方的生态型一般表现短日照类型。北方的生态型表现长日照类型。海洋性生态型要求较小的昼夜温差,大陆性生态型则要求较大的昼夜温差。南方的生态型种子发芽对低温春化没有明显要求,北方的生态型如不经低温春化,就不能打破休眠。在生化上,如乙醇酸氧化酶的活性也随气候类型(特别是温度)而异,大陆性生态型的酶活性随气温增加而加强的程度比海洋性生态型明显。这些生理反应都与其所在地区的气候特点有关。
(2)土壤生态型
主要是长期在不同土壤条件下分化形成的生态型。例如,牧草鸭茅生长在河滩地上和碎石堆上,由于土壤水分状况不同,形成两种不同的生态型。河滩上的牧草鸭茅,植株高大、叶肥厚、颜色绿、生长旺盛、刈草后易萌发、产量高;碎石堆上的牧草鸭茅,植株矮小、叶小、颜色较淡、萌生力极弱,产量也很低。在生理上也有差别,前者的细胞渗透压较后者为低。
(3)生物生态型
主要是在生物因子的作用下形成的生态型。例如同一种杨树的抗病虫能力不同。又如生活在不同植物群落中的稗,由于植物的竞争,在水稻田中茎秆直立,常与水稻同高,也差不多同时成熟,而生活在其他地方如牧场条件下则茎秆矮小。开花期迟早是两种不同的生物生态型。
4.1.1.3 他感作用
他感作用(allelopathy)也称异株克生,通常指一种植物通过向体外分泌代谢过程中的化学物质,对其他植物产生直接或间接的影响。这个概念是德国学者H . Molich于1937年提出的,已被大多数研究者所接受。这种作用是生存斗争的一种特殊形成,种内、种间关系都有此现象,如北美的黑胡桃,抑制离树干25m范围内植物的生长,彻底杀死许多植物,其根抽提物含有化学物质苯醌,可杀死紫花苜蓿和番茄类植物;香蒲发生种内竞争性异株克生,群丛中心枝叶枯萎;加利福尼亚灌木鼠尾草产生挥发性萜类.这种萜类为土壤黏粒所吸收,进入土壤表面,使土壤不适宜其他植物的生长。当湿润年度来临时或在火灾发生后,这种萜类物质大部分被冲洗或驱散,则1年生草类繁茂生长起来。
他感作用是进化过程中形成的一种普遍现象,它存在于各种气候条件下的各种群落中。他感作用中产生的化学物质以挥发气体的形式释放出来(这种情况多见于干旱地区)或者以水溶物的形式渗出、淋出或被分泌出来,它可能由地上部分或地下部分的活细胞释放,也可能来自它们的分解或腐烂以后。他感作用中植物的分泌物称为克生物质,对克生物质的提取、分离和鉴定已做了大量研究工作。
克生物质的毒害作用可能有不同方式,如抑制种子发芽,妨碍固氮菌的活动,或阻碍菌根的形成。例如帚石楠可使锡特云杉在英国造林失败,因为其克生物质可抑制云杉菌根的形成,因而栽植的云杉生长很慢,将帚石楠去除以后,云杉就能形成菌根,并且生长良好。蕨对很多树种的生长不利,这里既有对光、营养和水分的竞争的原因,也有由蕨渗出的酚酸对林木根系生长不利的原因。沙漠上的一些植物在遇到缺水胁迫时会产生自毒现象,从而利用有限的水分,避耐干旱,保持该物种的生存和繁衍。
他感作用具有重要的生态学意义:①对农林业生产和管理具有重要意义,如农林业的胁地现象就是由于他感作用使某些作物、果树不宜连作造成的。如早稻就是不宜连作的农作物,其根系分泌的对-羟基肉桂酸,对早稻幼苗起强烈的抑制作用,连作时则长势不好,产量下降;桃树根中发现扁桃苷,分解时产生苯甲醛,严重毒害桃树的更新生长。②他感作用对植物群落的种类组成有重要影响,是造成种类成分对群落的选择性以及某种植物的出现引起另一类消退的主要原因之一。如银胶菊原产于墨西哥,是一种产橡胶的草本植物,它群生时,不但本身生长不好,而且对周围植物产生很大的影响,这是因为银胶菊植物根系分泌出反肉桂酸,抑制自身及其他植物生长。③他感作用是引起植物群落演替的重要内在因素之。在美国东部阔叶林中的研究表明,黄桦(先锋树种)幼苗的根生长要受到糖槭(顶极群落树种)幼苗的抑制,这种抑制是由于槭树根尖分泌的一种物质造成的,在糖槭占据的地段,黄桦也很难更新,也可能是这种克生物质造成的。
他感作用对农业和林业的重要性日益显示出来。很多园艺种植者都知道烟草对很多观赏植物不利的现象。在农业中如何搭配作物,在林业生产中如何选择适宜的混交树种都有赖于对各物种之间他感作用的认识。
4.1.2 种间竞争
种间竞争(interspecific competition)是指两种或更多物种共同利用同样的有限资源时产生的相互竞争作用。当资源虽不短缺而两种发生彼此直接干扰时,亦可发生种间竞争。竞争的能力取决于种的生态习性、生活型、生态幅和生态位。近缘物种的植物种群,无论在对资源的需要和获取资源的手段上,竞争十分剧烈,特别是密度过大时更为激烈。生活型相同的不同种类的植物之间,也常常发生剧烈的竞争。物种处于其最适生态幅时,具有最大的竞争能力,例如,有些分布很广的种类,只有在它的最适生境处(即最能充分发挥其竞争能力的地方)才占优势。个体或物种的生态位(inche)(它所处的条件、利用资源和它在那种环境里发生的时间)是决定该个体或物种与其他个体或物种竞争程度的关键,一般竞争的程度越激烈,生态位重叠程度越大。此外,植物的生长速率、个体大小、抗逆性及叶子和根系的数目、生长习性(1年生还是多年生),以及禾本科、莎草科的植物产生分蘖的能力等,也都会影响竞争能力。
4.1.2.1 竞争的类型
一般可把竞争区分为干扰性竞争和利用性竞争两种类型。
(1)干扰性竞争(interference competition)
干扰性竞争是指竞争个体间直接的相互作用,即一种生物借助行为排斥另一种生物使其得不到资源。干扰竞争的最明显的例子是动物为了竞争领域或食物而进行的打斗。另外,他感作用也是一种典型的相互干扰性竞争。
(2)利用性竞争(exploitive competition)
利用性竞争是指竞争个体不直接相互作用,即指一种生物所利用的资源对另一种生物来说非常重要,亦即两种生物同时竞争利用同一种资源。例如在很多生境中,蚂蚁、啮齿类动物和鸟类都以植物种子为食。
竞争的结果可能是两个种群或多个种群形成协调的平衡状态,或者一个种群取代另一个种群;或有一个种群将另一个种群赶到别的空间中去,从而改变原生态系统的生物种群结构。
竞争结果的不对称性是种间竞争的一个共同特点。一个体的竞争代价常远高于另一个体。竞争杀死失败者是很普遍的,通过掠夺资源(使它们丧失资源)或通过干扰(直接杀伤或毒害它们),竞争不对称的例子大大超过对称性结果的例子。种间竞争的另一个共同特点是对一种资源的竞争能影响对另一种资源的竞争结果,如林分中林木间的竞争,林冠层中优势种减少了其他竞争者进行光合作用所需的太阳辐射能,这种对光的竞争也影响到根部吸收营养物质和水分的能力,也就是说,林木之间竞争中,根竞争与枝竞争之间有相互作用。
4.1.2.2 竞争排斥原理
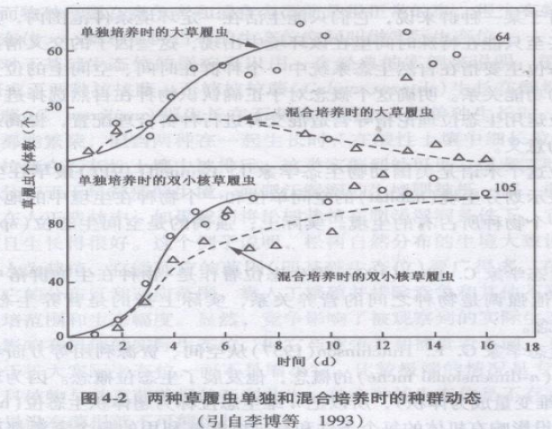
前苏联生态学家G .F . Gause(1934)首先用实验方法观察两个物种之间的竞争现象。他选择两种在分类上和生态习性上很接近的草履虫——双小核草履虫(Paramecium aurelia)和大草履虫(P .caudatum)进行实验。取两个种相等数目的个体,用同一种杆菌为饲料放在基本上恒定的环境里培养。开始时两个种都有增长,随后P .aurelia的个体数增加,而P .caudatum的个体数下降,16天后只有P .aurelia生存,而P .caudatum趋于灭亡(图4-2)。这两种草履虫之间没有分泌有害物质,主要是由于其中的一种增长快.而另一种增长慢,因竞争食物,增长快的物种排挤了增长慢的物种。其后,Park(1942,1954)用赤拟谷盗和杂拟谷盗混养所做的实验以及G .D .Tilman等(1981)用两种淡水硅藻和针杆藻所做的实验都得出了同样的结果。
这类实验说明:两个对同一资源产生竞争的种,不能长期在一起共存,最后导致一个种占优势,一个种被淘汰,这就是竞争排斥原理,或称之为Gause假说。
4.1.2.3 竞争的理论模型
美国学者Lotka(1925)和意大利学者Volterra(1926)分别独立地提出了描述种间竞争的模型,该模型是在Logistic方程的基础上建立起来的,它们具有共同的前提条件。
假定有两个物种,当它们单独生长时其增长形式符合逻辑斯谛模型,其增长方程是:

式中 N1,N2——分别为两个物种的种群数量;
K1,K2——分别为两个物种种群的环境容纳量;
r1,r2——分别为两个物种种群增长率。
如果将这两种放置在一起,则它们就要发生竞争,从而影响种群的增长。设物种1和物种2的竞争系数为α和β(α表示在物种l的环境中,每存在一个物种2的个体,对于物种1种群的效应;β表示在物种2的环境中,每存在一个物种1的个体,对于物种2种群的效应),并假定两种竞争之间的竞争系数保持稳定。则物种1、物种2在竞争中的种群增长方程为:
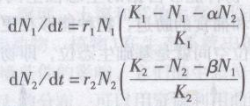
从理论上讲,两个物种竞争的结果是由两个种的竞争系数α、β、K1、K2比值的关系决定的,可能有以下4种结果:
(1)α>Kl/K2或β>K2/K1,两个物种都可能获胜;
(2)α> Kl/K2和β< K2/K1,物种l将被排斥,物种2取胜;
(3)α< Kl/K2和β> K2/K1,物种2将被排斥,物种1取胜;
(4)α< Kl/K2和β< K2/K1,两个物种共存,达到某种平衡。
高等植物种群混合栽培或培养时所表现出的竞争结果都可以用Lotka-Volterra竞争方程来说明。
4.1.2.4 生态位理论与应用
生态位是生态学中的一个重要概念,指物种在生物群落或生态系统中的地位和角色。对于某一种群来说,它们只能生活在一定环境条件范围内,并利用特定的资源。甚至只能在特殊时间里在该环境中出现,这些因子的交叉情况描述了生态位。生态位主要指在自然生态系统中一个种群在时间、空间上的位置及其相关种群之间的功能关系。明确这个概念对于正确认识物种在自然选择进化过程中的作用,以及运用生态位理论指导营造混交林进行种群合理配置,提高森林生产力具有重要的意义。
生态位这个术语是美国动物生态学家J .Grinnell(1917)最早在生态学中使用,用来表示划分生境(habitat)的空间单位和一个物种在生境中的地位。他认为生态位是一个物种所占有的生境。实际上,强调的是空间生态位(spatial niche)的概念。
英国生态学家C . Elton(1927)将生态位看作是“物种在生物群落中的地位与功能作用”他强调是物种之间的营养关系,实际上指的是营养生态位(trophic niche)的概念。
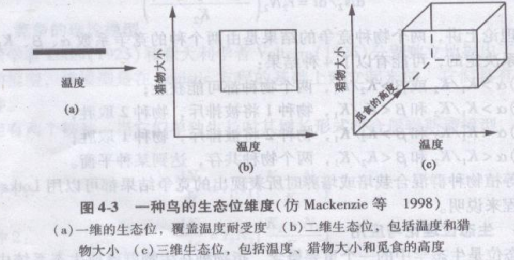
美国生态学家G . E . Hutchinson(1957))从空间、资源利用等方面考虑提出了n维生态位(n—dimensional niche)的概念,他发展了生态位概念。因为环境变量是多维的(三维变量成为体积),所以把n维生态位称为超体积生态位(hyper volume niche),假设影响有机体的每个条件和有机体能够利用的每个资源都可被当作一个轴或维,在此轴或维上,可以定义有机体将出现的一个范围,同时考虑一系列这样的维,就可以得到有机体生态位的一个增强了的定义图。举例说,苍头燕雀能耐受的温度范围与许多别的种互相重叠,然而考虑猎物大小和觅食高度是更多的维.就把苍头燕雀的生态位与其他许多种的生态位区分开来(图4-3)。n维超体积趣论在实践中有一个弱点,即不可能确定是否全部维都已经被考虑了,尽管如此,它是一个有用的概念。
另外,Hutchinson还提出了基础生态位(fundamental niche)与实际生态位(realized niche)的概念。一个物种能够占据的生态位空间,是受竞争和捕食强度影响。一般来说,没有竞争和捕食的胁迫,物种能够在更广的条件和资源范围得到繁荣,这种潜在的生态位空间就是基础生态位,即物种能栖息的理论上的最大空间。然而物种暴露在竞争者和捕食者面前是很正
常的事,很少有物种能全部占据基础生态位。一物种实际占有的生态位空间叫做实际生态位。
竞争对于基础生态位的影响可以用一个经典的实例来说明:植物生态学家Tansley研究丁两种拉拉藤:石楠拉拉藤生长在酸性土壤上,而细长拉拉藤则生长在石灰性土壤上。当单独生长时,两个种在两类土壤上都能繁荣,但当两种在一起生长时,在酸性土壤中细长拉拉藤被排斥,而石楠拉拉藤在石灰性土壤中被排斥,这类实例到处可见。很多二叶松在自然条件下一般分布于干旱贫瘠的生境,而肥沃湿润的生境则被栎、槭、椴等阔叶树占据,可是在人工造林中,如果我们将松树栽植在肥沃湿润条件下,它们也能正常生长,而且生长得很好。这个例子说明,松树自然分布的生境大致说明其实际生态位,而人为栽植,它能适应的范围(即基础生态位)要广得多,在自然界中每个种都有它的分布区和适应范围,靠人工繁殖并排除竞争和其他不利的关系,可扩大其栽培范围和生态幅度。显然,竞争影响了被观察到的实际生态位。另外互利共生也影响有机体的实际生态位,但它与竞争者和捕食者不同,互利共生者的存在倾向于扩大实际生态位,而不是缩小它。比较极端的情况是专性互利共生,如许多兰科植物与其真菌菌根的互利共生,单个种的生态位是不存在的,因为兰科植物如果没有菌根就不可能生长。
美国学者R .H . Whittaket(1970)认为,生态位是每个种在一定生境的群落中都有不同于其他种的自己的时间、空间位置,也包括在生物群落中的功能地位,并指出生态位的概念与生境和分布区的概念是不同的,生境是指生物生存的周围环境,分布区是指种分布的地理范围,生态位则说明在一个生物群落中某个种群的功能地位。
E .P . Odum(1971)将前人有关生态位的概念进行了综合,认为物种的生态位不仅决定于它们在那里生活,而且亦决定于它们如何生活以及如何受到其他生物的约束。生态位概念不仅包括生物占有的物理空间。还包括它在群落中的功能作用,以及它们在温度、湿度、土壤和其他生存条件的环境变化梯度中的位置。
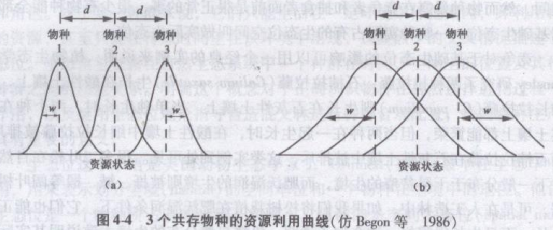
不同的生物物种(如动物、植物)在生态系统中的营养与功能的关系上各占据不同的地位,由于环境条件的影响,它们的生态位也会出现重叠与分化。不同生物在某一生态位维度上的分布,可以用资源利用曲线表示,该曲线常呈正态曲线(图4-4),表示物种具有的喜好位置及其散布在喜好位置周围的变异度。如图4-4(a)中各物种的生态位狭,相互重叠少,d>w,表示物种之间的种间竞争小;图4-4(b)中各物种的生态位宽,相互重叠多,d<w,表示种间竞争大。
比较两个或多个物种的资源利用曲线,就能全面分析生态位的重叠和分离情形,探讨竞争与进化的关系。如果两个物种的资源利用曲线完全分开,那么还有某些未被利用的资源。扩充利用范围的物种将在进化中获得好处;同时,生态位狭的物种,激烈的种内竞争更将促使其扩展资源的利用范围。由于这两个原因,进化将导致两物种的生态位靠近,重叠增加,种间竞争加剧。另一方面,生态位越接近,重叠越多,种间竞争就越激烈,将导致一物种灭亡或生态位分离。总之,种内竞争促使两物种生态位接近,种间竞争又促使两物种生态位分
离,这是两个相反的进化方向,那么.物种要共存,需要多少生态位分化呢?竞争物种在资源利用分化上的临界阈值称为相似性极限。在图4-4中d表示两物种在资源谱中的喜好位置之间的距离,w表示每一个物种在喜好位置周围的变异度,May等(1974)的分析结果表明,d/w=l可大致地作为相似性极限。
将前面讲述的竞争排斥原理与生态位概念应用到自然生物群落,则有以下一些要点: ’
(1)一个稳定的群落中占据了相同的生态位的两个物种,其中一个种终究要灭亡:
(2)一个稳定的群落中,由于各种群在群落中具有各自的生态位,种群间能避免直接的竞争,从而又保证了群落的稳定;
(3)一个相互起作用的、生态位分化的种群系统,各种群在它们对群落的时间、空间和资源的利用方面,以及相互作用的可能类型方面,都趋向于互相补充而不是直接竞争。因此,由多个种群组成的生物群落,要比单一种群的群落更能有效地利用环境资源,维持长期较高的生产力,具有更大的稳定性。
4.2捕食作用
捕食作用,即一种生物摄取其他种生物个体的全部或部分为食的关系。前者称为捕食者(predator),后者称为猎物或被食者(prey)。因此,这一广泛的定义包括:①“典型的捕食者”,在袭击猎物后迅速杀死而食之;②食草者,逐渐杀死对象生物(或不杀死)且只消费对象个体的一部分;③寄生者,与单一对象个体(寄主)有密切关系,通过寄生,生活在寄主的组织中。捕食者也可分为以植物组织为食的食草动物(herbivores),以动物组织为食的食肉动物(carnivores)以及以动植物两者为食的杂食动物(omnivores)。同时,两种类型的被捕食者都有保护自己的身体结构设置(如椰子或乌龟的厚壳)和对策,植物主要利用化学防御(chemical defenses),而动物则形成了一系列行为对策(behavioral strategies)。一方面,不同的捕食对策需要在不动的、具有化学防御性的被捕食者与能动而行为复杂的、但是美味的被捕食者之间进行权衡,从而在肉食者与草食者之间形成了进化趋异。另外,捕食者的食物变化很大,一些捕食者是食物选择性非常强的特化种(specialist),仅摄取一种类型的猎物;而另一些是泛化种(genera1ist),可吃几种类型的猎物。草食性动物一般比肉食性动物更加特化,或是吃一种类型食物的单食者(monophagous),或是以少数几种食物为食的寡食者(oligophagous),它们集中摄食具有相对防御性化学物质很少的几种植物。而草食性动物中的泛化种(或广食者,polyphagous)可通过避免取食毒性更大的部分或个体,而以一定范围的植物种类为食。动植物寄生者(parasite)都是特化种。
4.2.1 捕食者与猎物
4.2.1.1 捕食者与猎物的协同进化
捕食者与猎物的相互关系是经过长期的协同进化逐步形成的。为得到食物,捕食者必须首先搜寻猎物,然后捕食,所以捕食者形成了一整套适应性特征,如锐齿、利爪、毒腺或其他武器,诱惑追击,集体围猎,以提高捕食效率,以便更有力地捕食猎物。相反,猎物也形成了一系列行为对策,如保护色、警戒色、拟态、假死、集体抵御等以逃避被捕食。自然选择对于捕食者在于提高发现、捕获和取食猎物的效率,而对于猎物在于逃避、防止被捕食的效率,显然这两种选择是相互对立的。在捕食者与猎物的协同进化过程中,常会见到一种重要倾向,即有害的“负作用”倾向于减弱。在自然界中,捕食者将猎物种群捕食殆尽的事例很少。精明的捕食者大都不捕食已到繁殖年龄的猎物个体,因为这会降低猎物种群的生产力。被食者往往是猎物种群中老年或体弱患病、遗传特性较差的个体,捕食作用为猎物种群淘汰了劣质,从而防止了疾病的传播及不利的遗传因素的延续。人类利用生物资源,从某种意义上讲也要做“精明的捕食者”,不要过分消灭猎物,不然会导致许多生物资源灭绝。
捕食者与猎物的相互关系是生态系统长期进化过程中形成的复杂关系,它们是一对“孪生兄弟”,作为天敌的捕食者有时变成了猎物不可缺少的生存条件。不同生物种群之间存在的这种捕食者与猎物的关系,往往在对猎物种群的数量和质量上起着调节作用。例如1905年以前,美国亚利桑那州Kaibab草原的黑尾鹿种群保持在4000头左右的水平,这可能是美洲狮和狼的捕食作用产生的平衡,因为食物不形成限制因素。为了发展鹿群,政府有组织地捕猎美洲狮和狼群数量,鹿群开始上升,到1918年约40 000头,1925年,鹿群数量达到最高峰,约有10万头,但由于连续7年的过度利用,草场极度退化,鹿群的食物短缺,结果使鹿群数量猛降。这个例子说明,捕食者对猎物的种群数量起到了重要调节。
在自然环境中,捕食者与猎物的关系受许多因素的影响,往往是多种捕食和多种猎物交叉着发生联系。如果捕食者是多食性的,它就可以选择多种不同的食物,给自身带来更多的生存机会,也具有阻止被食者种群进一步下降的重要作用。相反,就被食者而言,当它的密度上升较高时,也会引来更多的捕食者,从而阻止其数量继续上升。例如,猫头鹰多以鹌鹑为食,当鹌鹑变少时,即转食啮齿类动物;在草原上,鼠类多的年份,以鼠为主要食物的黄鼠狼、狐狸、鹰等,有效地阻止鼠类种群数目的继续上升。
4.2.1.2 Lotka-Volterra捕食者~猎物模型
从理论上说,捕食者和被捕食者的种群数量变动是相关的,当捕食者密度增大时,被捕食者种群数量将下降;而当被捕食者数量降低到一定水平后,必然又会影响到捕食者的数量,随着捕食者密度的下降,捕食压力的减少,被捕食者种群又会再次增加,这样就形成了一个波动的种间数量动态(图4-5)。
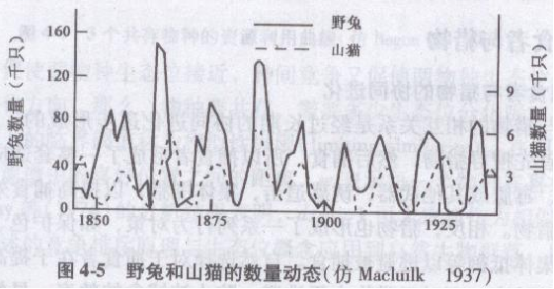
狭义概念的捕食者和被食者种群作用模型是Lotka(1925)和Volterra(1926)提出的,它的基本内容是:
对于被食者,可以假定在没有捕食的条件下,种群数量按几何级数增加:
![]()
式中 N——种群数量;
r2——被捕食者在没有捕食者时的瞬时增长率。
对于捕食者,可以假定在没有被食者的条件下,种群数量按几何级数减少:
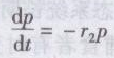
式中 p——捕食者密度;
r2——捕食者在没有教食者时的瞬时死亡率。
假定捕食者和被食者共存于一个有限的空间内,那么被食者的种群增长率因捕食而降低,其降低程度还随捕食者密度而变化,因此,被食者的种群方程可描述为:
dN/dt=(r1 -αp)N
同样,捕食者种群的增长率也将依赖于被食者的种群密度,捕食者种群方程可描述为:
dp/dt=(-r2+βN)p
式中,α是测度捕食压力的常数,即平均每一捕食者捕食猎物的常数。可以设想如果α=0,那么-αpN这一项等于零,那就表示被食者完全逃脱了捕食者的捕食。α值越大,就表示捕食者对于被食者的压力越大。因此,α可以被称为捕食压力常数。β是测度捕食者利用被食者而转化为更多的捕食者的效率常数。这个值越大,捕食效率越大,对于捕食者种群的增长的效应也就越大。因此,β可以称为捕食效率常数。
4.2.1.3 自然界中捕食者对猎物种群大小的影响
捕食者是否真能够调节其猎物种群的大小呢?目前有两种主要观点:
其一,有许多实例表明捕食者和食草动物对其猎物种群密度没有多大影响。主要因为:①任一捕食者的作用,只占猎物种群总死亡率的很小一部分,因此去除捕食者对猎物种群仅有微弱影响。如许多捕食者捕食田鼠,蛇仅是捕食者之一,所以去除蛇对田鼠种群数量影响不大。②捕食者只是利用了对象种群中超出环境所能支持的部分个体,所以对最终猎物种群大小没有影响。
其二,捕食者对猎物种群有致命性的影响。最有代表性的是向热带岛屿上引人捕食者后所导致的多种群灭绝。例如,太平洋关岛上引入林蛇后,有10种土著鸟消失或数量大大下降。存这些例子中猎物种群劣势很大,因为其没有被捕食的进化历史,也就段有发展相应的反捕食对策。然而,当猎物种群长期处于捕食者的捕获之下时,捕食的影响力也会很大。
4.2.1.4 猎物密度影响——功能反应
随着猎物密度的增加,捕食者可以捕获更多的猎物或可以较快地捕获猎物,直到最大,这种现象叫做捕食者的功能反应(functional response)。
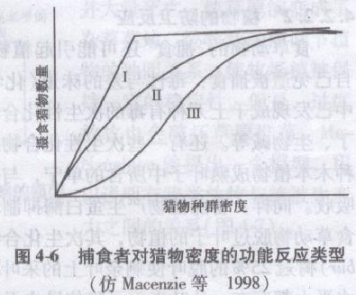 功能反应的概念最早被M .E . Solomon(1949)提出,后来又被C. C. Holling详尽地进行过研究。Holling提出了3种不同的功能反应类型(图4-6):I型功能反应是指每个捕食者所捕获的猎物数量随猎物密度的增加而呈线性增长,直达最大值为止。Ⅱ型功能反应是指每个捕食者所捕获的猎物是以递减的速度增加,直达最大值为止。Ⅲ型功能反应是指每个捕食者开始时所捕获的猎物很少,然后呈S形增长,并趋近于一个较高的渐近线。
功能反应的概念最早被M .E . Solomon(1949)提出,后来又被C. C. Holling详尽地进行过研究。Holling提出了3种不同的功能反应类型(图4-6):I型功能反应是指每个捕食者所捕获的猎物数量随猎物密度的增加而呈线性增长,直达最大值为止。Ⅱ型功能反应是指每个捕食者所捕获的猎物是以递减的速度增加,直达最大值为止。Ⅲ型功能反应是指每个捕食者开始时所捕获的猎物很少,然后呈S形增长,并趋近于一个较高的渐近线。
4.2.2食草作用
食草是广义捕食的一种类型。其特点是:①被食者只有部分机体受损害,通常捕食者只采食植物的某一部分,留下的部分能够再生;②植物本身没有逃脱食草动物的能力。
4.2.2.1 食草动物对植物的危害及植物的补偿作用
植物受食草动物捕食的危害程度随损害的部位、发育阶段的不同而异。在生长季早期.栎叶被损害会大大减少木材量,而在生长季晚期时,叶子受损害对木材产量可能影响不大。啮齿类动物和鸟类每年消耗大量乔灌草种子。在一定程度上控制着某些植物种群的数量。
有时食草动物与植物种群的数量变化,是一种激烈升降的变动。加拿大东部针叶林中生活的棕色卷蛾,一般每隔35~40年有一次大发生,棕色卷蛾食香脂冷杉的芽、花和针叶。大发生时,香脂冷杉的叶子被大量取食,林木濒于死亡,冷杉种群数量下降。如果只从周期性数量高峰来看,卷蛾是很多冷杉死亡的一种最大危害,但实际上,卷蛾的大发生与冷杉林的成熟有关,易受卷蛾危害的是成熟冷杉林分,幼树一般不易受卷蛾的攻击,老龄林木枯死后,林冠疏开,其冷杉幼树加速生长,很快到达主林冠层,因此自然条件下棕色卷蛾消长变化有助于森林更新。
林木的结实特性及种子和果实的大小、颜色等是长期协同进化的结果,也是对环境压力的适应表现,这包括对动物捕食的适应。大粒种子一般不是年年大量结实,存在结实间隔期,假若年年大量结实,捕食者会年复一年的依靠丰富食物增长种群数量,对树木来说将是灾难性的。
植物在被动物取食而受损害时,并不完全是被动的,而具有各种补偿机制。例如,植物的一些枝叶在受损害后,自然落叶减少而整株的光合效率可能加强。受害植物可能利用贮存于各组织和器官中的糖类得到补偿,或改变光合产物的分布,以维持根枝比的平衡。植物的受伤枝可能掉落,也可能将其糖类运输到未受伤害枝叶得到补偿。
4.2.2.2 植物的防卫反应
食草动物的“捕食”还可能引起植物的防卫反应,植物主要以两种方式保护自己免遭被捕食:毒性与差的味道(化学防御);防御结构(机械防御)。在植物中已发现成千上万种有毒的次生性化合物,如马利筋中的强心苷、烟草中的尼古丁、生物碱等。还有一些次生性化合物无毒,但会降低植物的食物价值。如许多种木本植物成熟叶子中所含的单宁,与蛋白质结合,使其难以被动物捕食者肠道吸收。同样,番茄植物产生蛋白酶抑制因子,可抑制草食者肠道中的蛋白酶。被食草动物脱过叶子的植物,其次生化合物水平会提高,例如欧洲白栎树冠25%的脱叶使剩余叶上的采叶蛾幼虫死亡率大大增加。防御结构在各种水平上都存在,从叶表面可陷住昆虫及其他无脊椎动物的微小绒毛(经常带钩或具有黏性分泌液)到大型钩、倒钩和刺,如荨麻、大蔷薇、冬青树和金合欢属(Acacia)植物阻止哺乳类食草动物。上述防御结构也可在脱叶的植物中被诱导出来,如被牛啃食后的悬钩子的皮刺较未啃食过的长而尖。
4.2.2.3 植物和食草动物的协同进化
在进化过程中,植物为防止食草动物的取食,发展了防卫机制,另一方面食草动物在进化过程中产生了相应的适应性,如形成特殊的酶进行解毒,出现了专性捕食者,如烟草天蛾不怕尼古丁,专食烟叶;澳大利亚的袋熊、袋貂以及许多昆虫都食有毒的桉叶。可见,植物保护剂只能抵制多食性捕食者,而不能抵制与被食者共同进化的专性捕食者
具有多种天敌的植物在进化中的一个重要趋势是逐渐成为只有一种天敌、一种专性捕食者的植物。对于植物来说,是一种选择有利性。多食性捕食者可以在根除了一种被食者后,再去捕食其他被食者,而专性捕食者不能做到这一点,当它们使被食者减少到一定程度以后,它们自己也会随之减少,这样又给植物以重新繁殖的机会。
共同进化中.物理的相互作用比化学的更加明显。可以普遍看到植物以小刺、软毛组织以及组织加厚等形式的机械防御的一些例子,足以说明植物与食草动物已发生的相互作用具有进化的长期性和关系的紧密性。
自然界亦存在相反的适应。植物能以各种方式取食昆虫和小动物,尽管这种食虫植物在自然界并不多见,全世界仅有500种左右,却代表着植物与动物协同进化的另一种形式。
4.2.2.4 植物与食草动物种群的相互动态
植物一食草动物系统也称为放牧系统(grazing system)。在放牧系统中,食草者与植物之间具有复杂的相互关系,简单认为食草动物的牧食会降低草场生产力是错误的。如在乌克兰草原上,曾保存500hm2的原始的针茅草原,禁止人们放牧。若干年后,那里长满杂草,变成不能放牧的地方,其原因是针茅的繁茂生长阻碍了其嫩枝发芽并大量死亡,使草原演变成了杂草草地。放牧活动能调节植物的种间关系,使牧场植被保持一定的稳定性。但是,过度放牧也会破坏草原群落。Mc-Naughton曾提出一个模型,用以说明有蹄类放牧与植被生产力之间的关系(图4-7)。
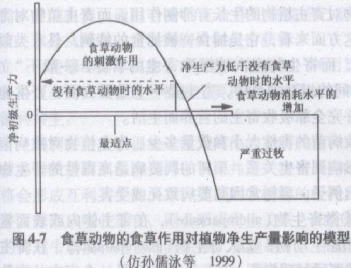 图4-7表明,在放牧系统中,食草动物的采食活动在一定范围内能刺激植物净生产力的提高,超过此范围净生产力开始降低,然后,随着放牧强度的增加,就会逐渐严重过度放牧的情形。该模型对牧场管理者具有重要意义。
图4-7表明,在放牧系统中,食草动物的采食活动在一定范围内能刺激植物净生产力的提高,超过此范围净生产力开始降低,然后,随着放牧强度的增加,就会逐渐严重过度放牧的情形。该模型对牧场管理者具有重要意义。
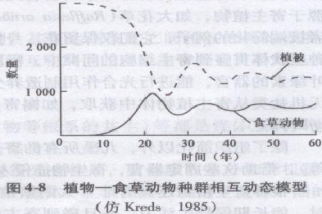 Caughley(1976)曾提出一个植物—食草动物相互作用放牧系统的种群相互动态模型,其基本思想与Lotka-Volterra的捕食者—猎物模型相同,但Lotka-Volterra是以指数增长描述猎物增长,而Caughley是以逻辑斯蒂方程描述植物种群增长。图4-8所模拟出的是植物和食草动物两个种群的相互动态过程。
Caughley(1976)曾提出一个植物—食草动物相互作用放牧系统的种群相互动态模型,其基本思想与Lotka-Volterra的捕食者—猎物模型相同,但Lotka-Volterra是以指数增长描述猎物增长,而Caughley是以逻辑斯蒂方程描述植物种群增长。图4-8所模拟出的是植物和食草动物两个种群的相互动态过程。
在一个生态系统中,捕食者与被捕食者一般保持着平衡,否则生态系统就不能存在,如果捕食者对猎物有害影响过分,则会导致一个和两个种群灭绝。尤其是具有两个负相互作用的种群.以往曾分别在不同的生态系统中,之后它们结合在一起时.有害作用就表现强烈,这是因为该系统对新来的生物还缺乏合适的调节机制或调节机制很弱所致。
4.3寄生与共生
4.3.1寄生
寄生是指一个种(寄生物)寄居于另一个种(寄主)的体内或体表,靠寄主体液、组织或已消化物质获取营养而生存。由于这样的营养关系,所以,寄生物使寄主植物的生长减弱,生物量和生产量下降,最后使寄主植物的养分耗竭。并使组织破坏而致死,因而,寄生物对寄主植物的生长有抑制作用,而寄主植物对寄生物则有加速生长的作用,从这方面来看,它是捕食一被捕食的特例,具有类似的性质。捕食者通常杀死猎物,而寄生物多次地摄取寄主的营养,一般不“立即”或直接杀死寄主。在植物之间的相互关系中,寄生是一个重要方面。寄生物以寄主的身体为定居的空间,并完全靠吸收寄主的营养而生活。
在寄生关系中,寄生物或致病菌的毒性大小和数量多少、寄主植物对致病菌的抗性强弱以及环境条件都会影响到寄生关系。栗树的凋萎病是高毒性的寄生物对一个无抗性的寄主植物的突出例子,栗树常因凋萎病致死或受害。
寄生物可以分为两大类:①微寄生物(microparasite),在寄主体内或表面繁殖,主要有病毒、细菌、真菌和原生动物:②大寄生物(macroparasite),在寄主体内或表面生长发育,但其增殖要通过感染期,从一个寄主到另一个寄主,多数在细胞间隙(植物)生长。营寄生生活的高等植物,可明显地分为全寄生和半寄生两类。全寄生植物含叶绿素的器官完全退化,无光合作用能力,因此营养全来源于寄主植物,如大花草、白粉藤属(Cissus)是有花植物寄生者极端简化的例子,它们仅保留花,身体所有其他器官都变为丝状的细胞束,这种丝状体贯穿到寄主细胞的间隙中,吸取寄主植物的营养。半寄生植物仅保留含叶绿素的器官,能进行光合作用制造养分,但根系发育不良或完全没有根,水和无机盐类从寄主植物体中获取,如槲寄生和小米草。
除了组织简化以外,几乎所有的寄生植物都出现专性固定器官(吸盘、小钩等),借助这些固定器官,寄生物能侵入并固定在寄主植物体内或体表。
很多寄生植物还具有非常大的繁殖能力和很强的生命力,在没有碰到寄主时,能长期保持生活力,一旦碰到寄主植物,又能立即恢复生长,营寄生生活。如寄生在很多禾本科植物根上的玄参科独脚金属(Striga)植物,一株可产生50万个种子,可保持生命力20年不发芽,但一旦碰到寄主植物时,其种子就开始发芽生长,并侵人和寄生在寄主根中。
多数的寄生植物只限于寄生在一定韵植物科、属中,即寄生具有一定的专性,这类寄生植物为专性寄生植物,如菟丝子属(Cuscuta)和列当属(Orobanche)中的很多种,常寄生在三叶草、亚麻、柳树、向日葵、大麻、苧麻等植物上。由于寄生具有一定的专性,所以寄生物和寄主常常是协同进化。
4.3.2共生
4.3.2.1 偏利共生
两个不同物种之间共生,对一方有利而对另一方无害的关系,称为偏利共生。
自然界中的附生现象,即一种植物定居在另一植物体的表呵,附生植物与被附生植物只在定居的空间上发生联系。它们之间没有营养物质的交流。如在热带和亚热带森林中,附生在树木上的地衣、苔藓、蕨类、兰花、仙人掌等植物,它们本身能自养,通过自身的光合作用制造自己所需的有机养料,所需的矿质元素、水分从降水、尘埃和腐烂树皮中获得。
一些攀缘藤本植物,本身茎不能直立,利用其他的树干作为机械支撑,从而获得更多的光照。这些植物主要存在于热带、亚热带潮湿的森林中,这也是一种偏利共生。
偏利共生关系很易过渡为其他的互利共生和寄生关系。如果附生植物产生的营养物质被雨水淋溶到树干下面并进入宿主周围的土壤中,被宿主吸收利用,则将会形成互利共生的关系。如果附生植物的根扎人树皮下面的韧皮部和木质部中并发育形成吸收器官,则会形成寄生关系,如在热带森林中,椿树可在一个宿主的树冠中萌发,用它的附生根,紧缠树干,并迅速长出气生根,到达地面变粗,结果使宿主生长受到严重的影响,同时榕树日益在宿主上面扩展,使宿主被遮荫,得不到充足的光照和养分,最后死亡,人们称热带森林中的这类植物为“绞杀植物”。
4.3.2.2 互利共生
两种生物生活在一起,两者相互有利,甚至达到彼此之间相互依赖的程度,这种现象称为互利共生。自然界中生物之间互利共生的现象非常普遍,形式也多种多样,概括来说主要有3类:
(1)连体互利共生
地衣、菌根、根瘤(固氮菌和豆科植物等根系的共生)等都是连体互利共生的典型例子,其两种生物长期接触,紧密结合在一起。
地衣是藻类和真菌的共生体,藻类进行光合作用,菌丝吸收水分和无机盐,两者结合,相互补充,共同形成一个统一的整体.生活在岩石或树干这样严酷的环境条件。
菌根是真菌和高等植物根系的共生体,真菌从高等植物根中吸收碳水化合物和其他有机化台物,或利用其根系分泌物,而又供给高等植物氮素和矿物质,二者互利共生。很多菌根植物在没有菌根时就不能正常生长或发芽,例如松树在设有与它共生的真菌的土壤里,吸收养分很少,以致生长缓慢乃至死亡。在缺乏相应真菌的土壤上造林或种植菌根植物时,可以在土壤内接种真菌,或使种子事先感染真菌,便能获得显著的效果。同样,某些真菌如不与一定种类的高等植物根系共生,也将不能存活。
在异养的固氮微生物和不能利用大气氮的自养植物之间存在着互利共生的关系。根瘤即是由于根瘤菌属(Rhizobium)的细菌侵人到豆科植物的根中形成的。"细菌由宿主处得到一个庇护所并且取得碳水化合物作为能量来源,而宿主植物则取得氯的来源,并因此可以在氮素缺乏的条件下生存。除豆科植物和根瘤菌属的共生外,放射线菌亦具与高等植物形成共生的特性。放射线菌类中的弗兰克氏菌属(Frankia)可侵人非豆科植物的根毛并形成根瘤,其固氮速度可等于或超过由根瘤菌在豆科植物上形成的根瘤。已知非豆科植物中有131属160种可有根瘤,其中在林业生产中晟重要的是赤杨和弗兰克氏菌属的关系,将赤杨作为混交树种提高林地生产力,在林业上日益受到重视。除根瘤外,还有叶瘤,这种现象常见于热带地区的植物。据不完全统计。叶瘤植物达370余种,茜草科的九节属(Psyvhotria)、茜木属(Paretta)、紫金牛科的硃砂根属(Ardisia)以及薯蓣科的薯蓣属(Dioscorea)等都是常见的叶瘤植物。
动物与微生物之间互利共生的例子很多,如反刍动物与其胃内的微生物间形成,一种互利共生的关系,微生物既帮助了反刍动物消化食物,自身又得到了生存。
(2)非连体互利共生
动物界有很多这种共生关系,两种生物不是长期结合在一起,而只有间断性的接触,蚂蚁和蚜虫之间的关系常受到人们的注意。另外,授粉是非连体互利共生的关系,大多数开花的双子叶植物,依靠传粉者(pollinator),可能是昆虫、鸟、蝙蝠或小型哺乳动物在植物间传递花粉。通常传粉者通过接受花蜜、油或花粉为食来获益。一些植物一传粉者的关系包含紧密的配对相互作用,两种互相依赖,如丝兰属植物与丝兰蛾科(Rodoxidea)昆虫以及无花果树与无花果寄生蜂之间所发生的关系。但是大多数植物一传粉者的关系比上述的更松散,每一传粉者用收获花蜜和花粉的植物都有一个范围,该范围在整个季节中随可获得的花的种类变化而改变。
除传粉,种子散布是另一类动物与植物之间的非连体共生。气流可非常有效地传播很小的种子,但大型种子仅能靠水流传播(如椰树种子)或靠动物散布。啮齿动物、蝙蝠、鸟类和蚂蚁都是重要的种子传播者。一些特化的种子传播者是种子采食者,它们摄食种子,但通过掉落、贮存或丢失种子帮助种子的散布,尽管这种种子丢失可能是偶然性的,但这种关系对双方还是相互有益的。另一些种子传播者包括食水果动物,它们摄食新鲜水果,但排除或去除种子,热带森林中75%的树种生产新鲜水果,其种子由动物散布。植物进化了这些富含能量的水果作为“报酬”来吸引食水果动物的取食,食水果动物一植物的关系通常是松散的,一些不同动物种类可取食同一种植物的水果。
(3)防御性互利共生
有一些互利共生为其中一方提供对捕食者或竞争者的防御。一些种类的草,如普通的多年生黑麦草与麦角真菌之间有互利共生关系,真菌生长在植物组织内或在叶子表面,生产具有很强毒性的植物碱。保护草免受食草者和食种子者的危害。蚂蚁一植物互利共生很普遍。许多植物在树干或叶子上有称做花外蜜腺的特化腺体,为蚂蚁提供食物源,该腺体分泌富含蛋白质和糖的液体。在许多种金合欢树中,蚂蚁也通过生活在树的空隙中得到物理保护。蚂蚁为其宿主提供对抗草食者很强的防御,并且有力地进攻任何入侵者。Janzen(1967)实验中把蚂蚁从金合欢树和Tachygali树上移走,这些树受到食草动物的取食水平大大提高,证明了蚂蚁的保护作用。
4.4种间协同进化
一个物种的进化达到成熟会改变作用于其他生物的选择压力,引起其他生物也发生变化,这种变化反过来又会引起相关物种的进一步变化,这种相互适应、相互作用的共生进化的关系即为种间协同进化(co-evolution)。
捕食者和猎物之间的相互作用可能是这种协同进化的最好实例。捕食对于捕食者和猎物都是一种强大的选择压力,捕食行为了生存必须获得狩猎的成功,而猎物为了生存则必须获得逃避捕食的能力。在捕食者的压力下,猎物必须靠增加隐蔽性、提高感官的敏锐和疾跑来减少被捕食的风险。所以,瞪羚为了不成为猎豹的牺牲品就会跑得越来越快,但瞪羚提高r了的奔跑速度反过来又成了作用于猎豹的一种选择压力,促使猎豹也增加奔跑速度。捕食者或猎物的每一点进步都会成为一种选择压力促进对方发生变化,即种间协同进化。
昆虫与植物之问的相互作用同捕食者与猎物之间的相互作用非常相似。、昆虫取食植物造成严重的损害,这对植物来说可能是一个强大的选择压力,如何逃避取食,对长命植物来说,由于更容易受到昆虫攻击,它们必须发展其他的防御方法。很多植物靠物理防御阻止具有刺吸式口器昆虫的攻击.如表皮加厚变得坚韧、多毛和生有棘刺等,还有一些植物则发展了化学舫御。 。
大型食草动物的啃食活动可对植物造成严重的损害,这无疑对植物也是一个强大的选择压力,在这种压力下,很多植物都采取了俯卧的生长方式或长得很高大。几乎所有的植物都靠增强再生能力和增加对营养生殖的依赖适应食草动物的啃食。最耐啃食的草本植物,其生长点都不在植物的顶尖而是基部,这样食草动物的啃食就不会影响它们的生长。
在寄生关系中,一种适应性很差的寄生物常遭到寄主的排除或致寄主于死命,在这两种情况下,寄生物都会死亡。相反,一种适应性很强的寄生物只带给寄主很小的损害,使寄主不仅可以忍受而且能较好地生存下去,这样寄生物本身的延续也就有了保障。寄生物最小的平衡状态,这种关系甚至会逐渐发展成一种互惠关系。
种间互利共生的协同进化可能发生在不同情况下,或来自寄生物寄主或捕食者一猎物之间的关系,或发生在没有协作或相互利益的紧密共栖者之间。例如,昆虫传粉可能起始于昆虫从风传播花上偷食花粉,然后双方的协同进化使双方从这种关系中获益。这样,在植物传粉者关系中,增强的传粉成功的优势产生出吸引昆虫的花(鲜艳的颜色、香味、花蜜),但是互利共生也可能“恶化”为一方对另一方利益非平衡的剥削——寄生。例如,许多兰花不为其传粉者提供任何奖励,而是通过气味、形状和色彩模式来模仿昆虫雌体以引诱昆虫落到花上(特别是蜜蜂和黄蜂),这是互利共生关系进化变为寄生关系的一个例子。